Evolution früher Landpflanzen
Reinhard Junker
 Die Fossilüberlieferung der ältesten Landpflanzen gilt allgemein als gute Stütze der Evolutionslehre. Der Autor zeigt, daß – im Detail betrachtet – eine Rekonstruktion evolutionärer Abfolgen jedoch kaum möglich ist. Die Formengruppen sind weniger stammbaumartig, sondern vielmehr netzartig miteinander verbunden, was im evolutionstheoretischen Deutungsrahmen kaum zu erwarten ist.
Die Fossilüberlieferung der ältesten Landpflanzen gilt allgemein als gute Stütze der Evolutionslehre. Der Autor zeigt, daß – im Detail betrachtet – eine Rekonstruktion evolutionärer Abfolgen jedoch kaum möglich ist. Die Formengruppen sind weniger stammbaumartig, sondern vielmehr netzartig miteinander verbunden, was im evolutionstheoretischen Deutungsrahmen kaum zu erwarten ist.
Eine bemerkenswerte Fallstudie zur Aussagekraft der Evolutionslehre im Bereich der Paläontologie.
Inhalt
| 1. | Gefäßpflanzen des Silurs und Unterdevons im Überblick | ||||||||||||||
| 1.1 | Silur und Unterdevon | ||||||||||||||
| 1.2 | Mittel- und Oberdevon | ||||||||||||||
| 2. | Argumente für eine evolutionäre Deutung der stratigraphischen Abfolge der Devonpflanzen im Überblick | ||||||||||||||
| 3. | Deutungsprobleme im Rahmen der Evolutionslehre im Überblick | ||||||||||||||
| 4. | Gibt es evolutionäre Übergangsformen unter den devonischen Landpflanzen? | ||||||||||||||
| Vorbemerkung: Landpflanzenreste im Silur | |||||||||||||||
| 4.1 | „Psilophyten“ | ||||||||||||||
|
|||||||||||||||
| 4.2 | Der evolutionäre Anschluß der unterdevonischen Lycopsiden | ||||||||||||||
| 4.3 | Taxa incertae sedis im Unterdevon | ||||||||||||||
| 4.4 | Systematische und evolutionäre Betrachtungen zu den Filicopsida (Pteropsida) des Devons | ||||||||||||||
|
|||||||||||||||
| 4.5 | Vorfahren der Sphenopsiden | ||||||||||||||
|
|||||||||||||||
| 4.6 | Progymnospermen | ||||||||||||||
|
|||||||||||||||
| 4.7 | Die baurnförmigen Lycopsiden des Devons | ||||||||||||||
|
|||||||||||||||
| 5. | Allgemeine evolutionäre Betrachtungen zu den devonischen Landpflanzen | ||||||||||||||
| 5.1 | Überblick | ||||||||||||||
| 5.2 | Gegenüberstellung von Deutungsansätzen | ||||||||||||||
| 5.3 | Die Deutung der Befunde im Einzelnen | ||||||||||||||
Literatur
Glossar
Artnamenverzeichnis
Vorbemerkungen
Lassen sich Fragen zum Ablauf der Geschichte des Lebens umso besser lösen, je mehr Fossilien ergraben werden? Der Ruf nach mehr Fossilien ist nicht ungewöhnlich. Doch nicht selten haben neue Fossilien vertraute Vorstellungen in Frage gestellt oder Probleme, die bereits gelöst schienen, neu aufgeworfen. Hin und wieder stellt sich nämlich im Nachhinein heraus, daß evolutionäre Szenarien deshalb stimmig rekonstruierbar waren, weil die Daten zu knapp und die Deutungsmöglichkeiten daher so vielfältig waren.
Die Geschichte der Entdeckung und der Deutung devonischer Landpflanzen kann diese Lektion der Paläontologie beispielhaft illustrieren. Die vorliegende Arbeit soll dies – eigentlich als Nebenergebnis – deutlich machen. In der Hauptsache geht es um eine Vorstellung der eigentümlichen Welt der Landpflanzen des Devons, einer Zeit, in der nach evolutionstheoretischen Vorstellungen die Radiation der wichtigsten Landpflanzengruppen stattfand. Die Bedeutung der Thematik für die Paläontologie und Evolutionsbiologie ist damit umrissen.
Inwieweit lassen sich aber aufgrund der zu Tage geförderten Überreste dieser Pflanzen früherer Zeiten evolutionäre Abstammungszusammenhänge begründen? Dies ist die Leitfrage, unter der an die Materie herangegangen wurde. Es sollte versucht werden, die in der Literatur beschriebenen fossilen Funde und vorgelegten evolutionstheoretischen Deutungen auf diese Frage hin zu untersuchen. Die Darstellung beschränkt sich im wesentlichen auf das Devon. Auf silurische und karbonische Pflanzengruppen wird nur eingegangen, wenn sie zum Verständnis von Problemen der devonischen Pflanzenwelt wichtig sind. Ebenso bleibt die Frage ausgeklammert, woher die devonischen Pflanzen kommen – eine Frage, zu der Fossilien nach einhelliger Auskunft der Paläobotaniker ohnehin nichts Wesentliches beitragen können. Zweifellos sind diese Beschränkungen an manchen Stellen etwas künstlich; sie haben aber den Vorteil, daß die Darstellung überschaubar bleibt.
Die neuerdings auch in der Botanik an Bedeutung zunehmende Cladistik wurde ebenfalls nicht berücksichtigt. Angesichts der vielfach fragmentarischen Natur der Fossilüberlieferung devonischer Pflanzen und der geringen Bedeutung, die den Fossilien in der Cladistik beigemessen werden, ist dies zu rechtfertigen.
Abschließend noch ein praktischer Hinweis: Dem nicht mit der Materie vertrauten Leser wird Kapitel 4, in dem die relevanten Daten im Detail nach dem System der Pflanzen und stratigraphisch geordnet präsentiert werden, stellenweise zu speziell sein. Die wesentlichen Daten und Schlußfolgerungen werden jedoch im 5. Kapitel aufgegriffen und nach thematischer Ordnung diskutiert. Durch die dort angebrachten Querverweise und mit Hilfe des Artnamenverzeichnisses kann gezielt auf die ausführlichere Dokumentation in Kapitel 4 zurückgegriffen werden. Auf diese Weise – so die Hoffnung des Autors – kann nicht nur der Spezialist, sondern auch jeder an Fragen der Evolutionslehre interessierte Biologe Zugang zu diesem interessanten Feld der Paläobotanik finden.
Reinhard Junker, Mai 1996
Zusammenfassung
Die stratigraphisch ältesten Landpflanzengruppen des Silurs und des Devons werden vorgestellt. Es wird der Frage nachgegangen, inwieweit die Fossilüberlieferung die Vorstellung evolutionärer Abfolgen stützt. Die ältesten unumstrittenen gefäßführenden Landpflanzen treten im Obersilur auf; die stratigraphische Stellung einiger silurischer Gefäßpflanzenreste ist umstritten; die Identifikation oder die Gefäßpflanzennatur älterer silurischer Funde ist unsicher. Im Laufe des Devons ist eine deutliche Komplexitätszunahme der fossil überlieferten Formen festzustellen. Die Rekonstruktion evolutionärer Abfolgen erweist sich im Detail jedoch als schwierig oder unmöglich. Zum Teil fehlen überzeugende Zwischenformen. Der Mosaikcharakter vieler Gattungen und zahlreiche Konvergenzen verhindern in vielen Fällen eine widerspruchsfreie Einordnung in evolutionäre Reihen. Die einzelnen Gattungen oder supragenerischen Taxa sind vielfach netzartig durch Merkmalsübereinstimmungen verbunden. Wenn einigermaßen widerspruchsfreie evolutionäre Reihen konstruiert werden können, ist die zugrundeliegende Datenbasis eher schmal. Einige Pflanzenreste sind weder taxonomisch noch evolutionär einzuordnen. Innerhalb der taxonomischen Gruppen kann in der Regel keine mit der Stratigraphie einhergehende evolutionäre Tendenz festgestellt werden.
5. Allgemeine evolutionäre Betrachtungen zu den devonischen Landpflanzen
5.1 Überblick
Läßt man die Vielfalt der Pflanzengestalten des Devons Revue passieren, so entsteht der Eindruck einer ziemlich kunterbunten Kombination von Einzelmerkmalen. In der Stelenanatomie, der Verzweigung, der Anordnung und dem Bau der Sporangien können zwar bei Einzelbetrachtungen grobe Tendenzen festgestellt werden, doch im Bereich der feineren Systematik (auf dem Niveau von Ordnungen, Familien und Gattungen) steht die Ableitung evolutionärer Abfolgen aufgrund der unterschiedlichen mosaikartigen Kombination von Pflanzenstrukturen großen Schwierigkeiten gegenüber. Dies äußert sich in folgenden Beobachtungen:
Je nach Merkmalsgewichtung ergeben sich unterschiedliche Möglichkeiten, Gattungen zu Familien oder Ordnungen zusammenzufassen; die Systematik ist im Fluß und konnte bisher in vielen Fällen nicht gefestigt werden (vgl. Tab. 6). 1964 schrieb Banks, „the grouping of genera into families and orders is more a matter of convenience than of proven natural affinity“ (S. 15f.). Diese Situation hat sich nicht wesentlich geändert.
- einzelne Gattungen weisen zu vielen unterschiedlichen Taxa Gemeinsamkeiten auf (Abb. 48),
- es gibt sozusagen zu viele Zwischenformen, d. h. die einzelnen Gattungen sind durch Merkmalsübereinstimmungen netzartig miteinander verbunden (Abb. 49). Beispielhaft für diese Situation sind die Zosterophyllophyten. Kenrick & Edwards (1988, 111) sind bezüglich dieser Gruppe der Auffassung, daß die sich überlappende Verkettung der Merkmale weitere Unterteilungen der Zosterophyllophyten, z. B. in Familien, nach dem gegenwärtigen Kenntnißtand unhaltbar machen.
- viele devonische Pflanzenordnungen weisen nur wenige Gattungen auf,
- Stammbäume können nur unter der Annahme zahlreicher Konvergenzen oder Reversionen konstruiert werden (vgl. Gensel 1992, 450).
- z. T. ist die Bestimmung von Homologien schwierig (vgl. beispielhaft Gensel 1992, 450; Fig. 12).
Dazu kommen weitere Befunde, die evolutionstheoretische Deutungen erschweren:
- die nach der Merkmalsverteilung passendsten Stammbaumrekonstruktionen stehen mit der stratigraphischen Abfolge oft nicht in Einklang; mögliche Zwischenformen treten regelmäßig stratigraphisch zu spät auf (Tab. 11, S. 73),
- innerhalb engerer Verwandtschaftskreise sind Abfolgen in Richtung größerer Komplexität stratigraphisch oft nicht festzustellen, Merkmalsreihen entsprechen vielfach nicht der Stratigraphie.
- manche Pflanzengruppen stehen ziemlich isoliert (Pseudoborniales, einige Taxa incertae sedis).
Wie gut ist demnach eine evolutionäre Deutung der Entfaltung der devonischen Landpflanzenwelt begründet? In der Beantwortung dieser Grundfrage lassen sich subjektive Wertungen nicht vermeiden. Nachfolgend wird eine evolutionskritische Perspektive eingenommen. Der Befürworter der Evolutionslehre hat die Möglichkeit, gezielt gegen die vorgetragenen Interpretationen zu argumentieren. Zuvor sollen unterschiedliche Deutungsansätze kurz umrissen werden.
Abb. 48: Merkmalsbeziehungen unter devonischen Landpflanzengruppen.

A nach Taylor & Taylor 1993, 361; Stewart & Rothwell 1993, 216;
B nach Skog & Banks 1973, Stein et al. 1983, Gensel & Andrews 1984, Meyen 1987
C nach Taylor & Taylor 1993, 363
D nach Taylor & Taylor 1993, 367f.; Scheckler 1974, 468, 470
E nach Andrews et al. 1974
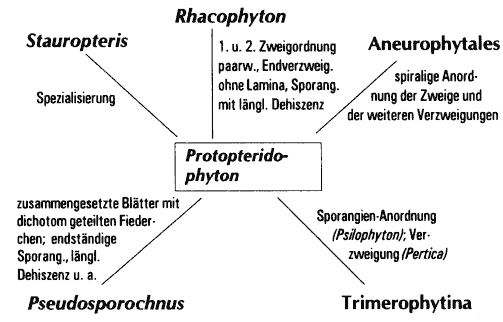
F nach Taylor & Taylor 1993, 366, Li & Hsü 1987
5.2 Gegenüberstellung von Deutungsansätzen
Als Gründe für unterschiedliche Fossilinhalte in aufeinanderfolgenden Sedimentgesteinen kommen in Frage:
1. Evolution. Das entspricht dem gängigen Paradigma. Das zeitlich gestaffelte Auftreten unterschiedlicher Pflanzentypen wird außer in Ausnahmefällen auf eine zeitlich gestaffelte evolutionäre Entstehung zurückgeführt.
2. Ökologische Gründe.
a. Durch eine Neu- oder Wiederbesiedlung brachliegender Flächen während kurzer Zeiträume kann es ebenfalls zu einem zeitlich abgestuften Auftreten unterschiedlicher, in der Komplexität zunehmender Pflanzengemeinschaften kommen. Eine Besiedlung heutiger Brachländer würde jedenfalls auch eine schrittweise Komplexitätszunahme der sich einfindenden Pflanzen aufweisen.
b. Die fossil konservierten Pflanzengemeinschaften könnten Ökosystemen mit unterschiedlichem Komplexitätsgrad entsprechen, die aus geologischen Gründen zu unterschiedlichen Zeiten zur Ablagerung gelangten. Auch hier muß ein Kurzzeitszenario zugrundegelegt werden. Eine solche Erklärung müßte plausibel machen, wie es zur Konservierung der vorgefundenen Abfolge kommen konnte. Auch heute können Ökosysteme unterschiedlichen Komplexitätsgrads unterschieden werden.
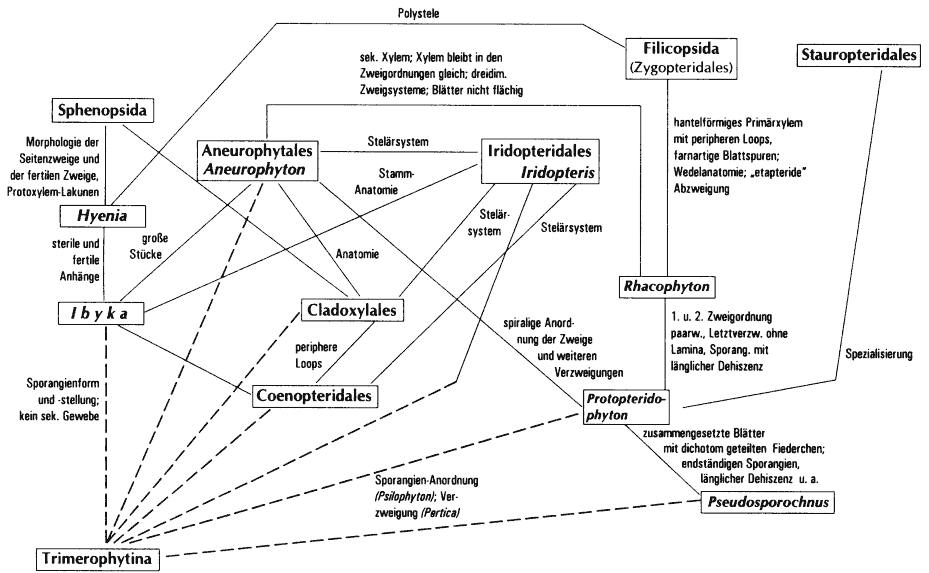
Abb. 49: Synopse der Merkmalsberzeichnungen unter devonischen Landpflanzengruppen aus Abb. 48
5.3 Die Deutung der Befunde im Einzelnen
5.3.1 Komplexitätszunahme
Wie bereits in der anfangs vorgelegten Übersicht (Kapitel 2) erwähnt, ist ein Hauptargument für eine evolutionstheoretische Deutung die grobe Tendenz zur Komplexitätszunahme im Laufe des Devons und die Tatsache, daß die ältesten Landpflanzenreste auch die strukturell einfachsten sind. Wie oben kurz umrissen, kommen auch ökologische Ursachen als Deutung dieses Befundes in Frage (Besiedlungsphasen, Konservierung unterschiedlicher Biotope in unterschiedlichen Zeiten).
Innerhalb engerer Verwandtschaftskreise ist bei den Deutungen Vorsicht geboten: Variabilität von Arten kann angesichts der fragmentarischen Überlieferung evolutionäre Tendenzen vortäuschen. So können beispielweise bei Huvenia aus den Sporangienträgern an einer einzigen Pflanze morphologische Reihen konstruiert werden, die von verzweigten Achsen bis zu fast vollständig reduzierten Achsen reicht (Hass & Remy 1991, 158). Dadurch wird die Brauchbarkeit dieses Merkmals für phylogenetische Betrachtungen relativiert.
5.3.2 Stammbaumdarstellungen
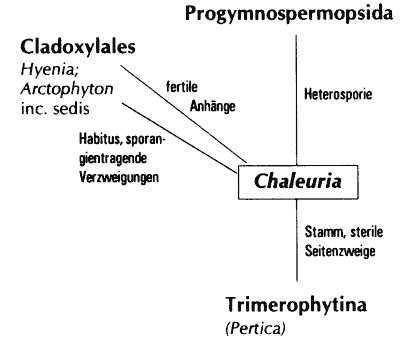
Bei vergröberter Betrachtung können die unterdevonischen Pflanzengruppen in einem Stammbaum angeordnet werden. Dabei bleibt der Anschluß einiger Gattungen und höherer Taxa fraglich (Chaleuria, Weylandia, Duisbergia, die Sphenopsiden-Ordnungen, Barinophytales, Paläophyllales) oder es bestehen konkurrierende Theorien zu evolutionären Anschlüssen. Betrachtet man jedoch die Formenvielfalt devonischer Pflanzen auf taxonomisch niederen Niveaus, ist es kaum noch möglich, Abstammungsverhältnisse zu klären. Das hat verschiedene Gründe, vor allem der Mosaikcharakter der Merkmalskomplexe vieler Taxa und damit verbunden das Auftreten zahlreicher Konvergenzen und Heterobathmien (s. u.). Stein et al. (1984) haben das beispielhaft aufgezeigt (Abschnitt 4.5.3). Erschwerend kommt hinzu, daß viele Gattungen nur bruchstückhaft bekannt sind. Wenn nun aber schon die bruchstückhafte Dokumentation fossiler Devonpflanzen enorme Schwierigkeiten in der Taxonomie und der phylogenetischen Rekonstruktion bereitet, ist durch weitere Zunahme der Formenkenntnis eine Besserung der Situation kaum zu erwarten. Denn bereits offenkundige Merkmalswidersprüche können dadurch nicht beseitigt werden. Vielmehr muß damit gerechnet werden, daß die Situation noch komplizierter wird. Aber das wird die Zukunft erweisen müssen.
5.3.3 Evolutionäre Reihen
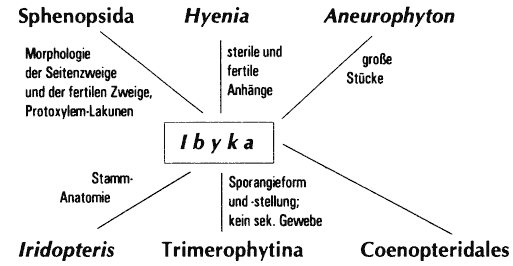
Wie vorgeführt, können in Teilbereichen evolutionäre Reihen konstruiert werden, wobei in der Regel idealisierend vorgegangen werden muß: Sehr häufig passen Übergangsformen stratigraphisch nicht (vgl. Abschnitte 4.1.1 [z.B. Aglaophyton], 4.2 [z. B. Asteroxylon], 4.4.1 [Rhymokalon], 4.5.3 [z. B. Ibyka] oder 4.6.2 [Aneurophytales]). Evolutionäre Reihen besitzen daher oft nur Modellcharakter.
Soweit evolutionäre Reihen aufgrund der Merkmalsverteilung und der Stratigraphie einigermaßen plausibel gemacht werden können, ist die „Beweiskraft“ der zugrundeliegenden Indizien oft relativ schwach. So können zwar die Trimerophyten durchaus als evolutionäre Vorstufen zu mitteldevonischen Pflanzengruppen angesehen werden, doch ist diese Gruppe so undifferenziert, daß fast alle mitteldevonischen Pflanzengruppen auf sie zurückgeführt werden können (Abb. 50). Die Merkmalsübereinstimmungen betreffen hauptsächlich (oder ausschließlich) sogenannte Primitivmerkmale, die nicht sehr aussagekräftig sind (Beispiel: Die evolutionäre Rolle des Trimerophyten Psilophyton in der Diskussion von Banks et al. 1975, 112f.). Auch an das Beispiel Oocampsa (Abschnitt 4.6.1) sei hier erinnert, eine als evolutionäre Zwischenform vergleichsweise gut interpretierbare Gattung, von der aber wichtige Merkmale unbekannt sind, so daß eine evolutionäre Zwischenstellung eigentlich nur auf vorläufigen Verdacht konstatiert werden kann.
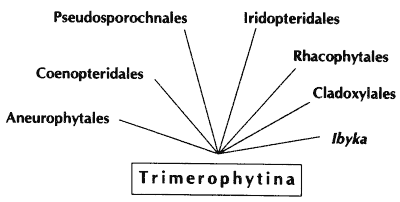
Abb. 50: Die Trimerophyten werden als Ausgangsgruppe für zahlreiche mittel- und oberdevonische Pflanzengruppen diskutiert.
5.3.4 Übergangsformen bei einzelnen Merkmalen bzw. Merkmalskomplexen und konstruktionsmorphologische Gesichtspunkte
Das zahlreiche Vorkommen unterschiedlich abgestufter Merkmalsausprägungen (z. B. unterschiedlich starke Lobierung der Stele, Flächigwerden dreidimensionaler Astsysteme, Anordnung der Sporangien, Übergänge zwischen Homo- und Heterosporie) kann als Ausdruck evolutionärer Prozesse gewertet werden. Im Rahmen einer evolutionstheoretischen Interpretation sollten die eingetretenen Veränderungen durch „Fitneßgewinn“ und unter Berücksichtigung konstruktionsmorphologischer Gesichtspunkte verständlich gemacht werden. Das heißt: Zum einen sollte verständlich gemacht werden, weshalb die mutmaßlichen evolutionären Übergänge Überlebensvorteile beinhalten, zum anderen, inwiefern die konstruktiven Vorbedingungen zu den jeweiligen Folgekonstruktionen führen konnten oder führen mußten. „There are constraints dü to the historical or phylogenetic burden of organisms frequently leading to parallelisms“ (Mosbrugger 1992, 20). Diese Gesichtspunkte scheinen in der paläobotanischen Literatur über die Devonpflanzen unterrepräsentiert zu sein.
Ein interessantes Beispiel diskutiert Mosbrugger (1992): Die Entstehung von Mikrophyllen kann nicht einfach unter dem Gesichtspunkt des Fitneßgewinns gesehen werden. Denn überraschenderweise ist das Verhältnis Oberfläche zu Volumen bei den beblätterten Gattungen Asteroxylon und Drepanophycus ungünstiger als bei der unbeblätterten Gattung Aglaophyton. Der Grund liegt im größeren Durchmesser der Achsen der ersteren. Damit entfällt das Argument, die Bildung von Mikrophyllen sei eine Verbesserung; in Wirklichkeit war sie einfach konstruktiv notwendig, damit die Versorgung bei größerem Achsendurchmesser gewährleistet bleibt.
Eine evolutionäre Deutung für das Auftreten unterschiedlicher Konstruktionen ist nur dann zwingend oder naheliegend, wenn nachgewiesen werden könnte, daß gewisse Übergangsstadien (z. B. unterschiedliches Ausmaß von Flächigwerden von Astsystemen) nicht durch funktionelle Erfordernisse erklärt werden könnten. Dieser Nachweis ist aber beliebig schwierig. Denn wie will man begründen, daß ein teilweise planiertes Astsystem nicht die optimale Ausprägung für die betreffende Pflanze in ihrem speziellen Lebensraum darstellt? Man kann nur den Verdacht hegen, daß es so sein könnte. Dazu kommt die Schwierigkeit, daß die devonischen Pflanzen oft nur unvollständig erhalten und die Lebensräume ungenügend bekannt ist. Ohne ausreichende Kenntnis der Lebensbedingungen kann aber die funktionelle Bedeutung organismischer Strukturen kaum beurteilt werden.
Man kann noch weiter fragen, um beim Beispiel der Planation zu bleiben, weshalb eine teilweise Planation von Sproßsystemen erfolgen soll, wenn sie keinen adaptiven Wert hat. Was war der Motor der teilweisen Planation, wenn sie keinen Vorteil brächte? Es ist also anzunehmen, daß es einen guten Sinn hatte, daß in der Vergangenheit Pflanzen mit teilweise planierten Zweigsystemen existierten. Vielleicht ist es aufgrund mangelhafter Kenntnisse über diese Pflanzen (noch) nicht möglich, diese Ausprägung funktionell zu verstehen.
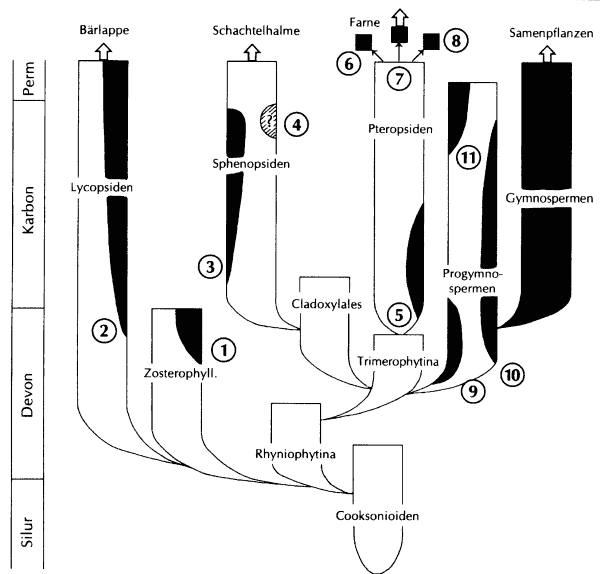
Abb. 51: Überblick über das unabhängige Auftreten von Heterosporie (schwarze Ausschnitte im Stammbaum). Die evolutionären Verbindungen und die Monophylie der angegebenen Taxa sind oft unsicher. 1 Einige Barinophytales, 2 Selaginellales und Rhizomorphales, 3 einige Equisetales, 4 eventuell einige Sphenophyllales, 5 einige Stauropteridaales, 6 Salviniales, 7 Marsileiales, 8 einige Filicales, 9 einige Aneurophytales, 10 einige Archaeopteridalesm 11 einige Noeggerathiales (nach Bateman & DiMichele 1994)
5.3.5 Konvergenzen und Heterobathmien
Aufgrund der im Vergleich zu Tieren offeneren Konstruktion der Pflanzen sind weniger „constraints“ bei evolutionären Veränderungen zu erwarten. So scheint beispielsweise die Anordnung von Sporangien konstruktiv unabhängig von der Stammanatomie zu sein. Die devonischen Landpflanzen zeichnen sich durch das zahlreiche Auftreten von Heterobathmien aus: ursprüngliche und abgeleitete Merkmale sind bei vielen Taxa in bunter Mischung anzutreffen (Tab. 9). „Many possible combinations of anatomy and morphology are becoming evident among early land plants“ (Gensel 1992, 451). Oft sind Anatomie und Morphologie unterschiedlich zu bewerten: „The morphology of the Devonian forms may suggest one affinity, their anatomy another“ (Skog & Banks 1973, 376).
|
|
||
|
|
||
| Pseudobornia | Bau der Stele | Blattstellung |
| Aneurophytales | Verzweigung | Holzanatomie |
| Uskiella (Rhynioph.) | vegetative Teile | Sporangienbau |
| Leelercqia (Lycops.) | gabelig verzweigte Mikrophylle, Protostele, Homosporie | Ligula, Art der tracheidalen Wandverstärkung |
| Sphenophyllales/ Calamitales |
gleich im fertilen Bereich, unterschiedlich im Xylem (Schweitzer 1983, 101) | |
| Archaeocalamites (Equisetales) | Sterile Anhänge in Wirteln, mehrfach dichotom gegabelt, keinerlei Verwachsung | Sekundärholztracheiden besitzen moderne („sehr fortschrittliche“) Hoftüpfel; Eustele mit Carinalkanälen |
|
|
||
Im evolutionstheoretischen Deutungsrahmen kommt man nicht umhin, zahlreiche Konvergenzen zu postulieren. So muß beispielsweise angenommen werden, daß Heterosporie im Devon und Karbon mindestens 11mal (vielleicht bis zu 20 mal) unabhängig aufgetreten ist (Bateman & DiMichele 1994; Abb. 51). Ein weiteres bemerkenswertes Beispiel ist das Auftreten von Gefäßbahnen mit peripheren Loops, die z. T. aus Parenchym, z. T. aus Protoxylem bestehen, bei Pflanzen sehr unterschiedlicher Affinität. In Tab. 10 sind die wichtigsten Beispiele zusammengestellt.
- Heterosporie (in mindestens 11 Taxa unabhängig aufgetreten)
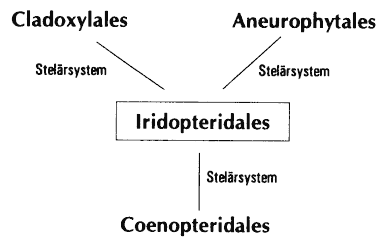
- Periphere Loops (z. T. Cladoxylales, Rhacophyton, z. T. Iridopteridales)
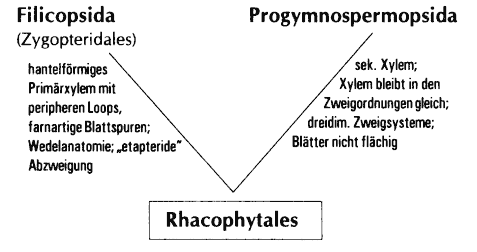
- Sekundärxylem (Progymnospermen, z. T. Cladoxylales, Zygopteris, Archaeocalamites, Duisbergia, Rhacophyton)
- Flächige Blätter (Archaeopteridales, Filicopsida, Pteridospermae)
- Angular-Organe (z. T. Zosterophyllophytina, z. T. Trimerophytina; vergleichbare Organe bei einigen Rhyniophytina; vgl. S. 32)
- Mikrophylle? (vgl. Diskussion in Abschnitt 4.2) Tracheiden (unsicher)
- Sporangienteilung in zwei Hälften (Renalia, Hsuea, einige Zosterophyllophyten?) (Edwards & Edwards 1986, 215; Shute & Edwards 1989, 133).
- ähnliche Tracheidenwandverstärkungen bei einigen Rhyniophyten, Zosterophyllophyten und Trimerophyten (Taylor 1986, 82).
Das oftmalige unabhängige Auftreten von Heterosporie macht entsprechende Selektionswirkungen wahrscheinlich (Bateman & DiMichele 1994, 406). Auffallend ist, daß fast alle heterosporen Linien in der relativ kurzen Periode des Oberdevons und Karbons auftraten (Bateman & DiMichele 1994, 407). Spielen ökologische Gründe dafür eine maßgebliche Rolle?
Die große Zahl von Konvergenzen ist evolutionstheoretisch nicht leicht zu deuten. Weshalb, so muß man fragen, führten – darwinistisch argumentiert – ähnliche Selektionsdrücke bzw. – konstruktionsmorphologisch argumentiert – bestimmte Vorkonstruktionen – so häufig zu ähnlichen Folgestrukturen?
5.3.6 Taxa incertae sedis
Die taxonomisch umstrittenen bzw. nicht einordenbaren Taxa (vgl. Abschnitte 4.3, 4.6.6, 4.7.2 und einige in Abschnitt 4.4 erwähnte Gattungen) stellen ebenfalls eine Herausforderung für evolutionstheoretische Betrachtungen dar. Die Deutung, es handle sich dabei um Zeugnisse einer experimentellen Phase (s. Abschnitt 5.3.11) der Evolution, ist zunächst nur eine Beschreibung des Befundes ohne Erklärung. Es muß erklärt werden, weshalb in einer solchen Phase besonders viele Konvergenzen auftreten.
5.3.7 Geringe Artenzahlen in Ordnungen
Wenn man die Taxonomie heutiger Pflanzengruppen, insbesondere der Blütenpflanzen mit der Taxonomie devonischer Pflanzen vergleicht, so fällt die häufig anzutreffende geringe Zahl von Arten und Gattungen je Ordnung, Unterklasse oder Klasse auf (vgl. Tab. 6, S. 55). So wurden zu den Trimerophyten bislang nur 3 oder 4 Gattungen gestellt (abgesehen von Formgattungen, die wahrscheinlich zu den anderen Taxa gerechnet werden müssen), die Iridopteridales weisen ebenfalls kaum eine Handvoll Gattungen auf, ebenso die Aneurophytales, die Archäopteridales und die Stauropteridales. Die Pseudoborniales sind monotypisch. Auch bei den Barinophytales werden nur zwei Gattungen unterschieden. Dazu kommen noch Taxa, deren Existenz aber von den meisten Autoren nicht anerkannt wird, wie etwa die Ibykales (vgl. Abschnitt 4.5.2) oder die Drepanophycopsida (vgl. Abschnitt 4.2). Wie ist dieser Befund zu werten? An folgende Deutungsmöglichkeiten kann gedacht werden: Zum einen könnte diese Situation auf eine große Lückenhaftigkeit der Fossilüberlieferung hinweisen. Oder die Aufstellung neuer Taxa erfolgt zu großzügig; vielleicht werden manche Merkmalsunterschiede zu sehr gewichtet und die Variabilität der Arten, Gattungen oder Familien unterschätzt. Schließlich kann diese Situation aber auch evolutionskritisch als Hinweis dafür gewertet werden, daß evolutionäre Zusammenhänge gar nicht vorhanden sind und wir es mit diskreten Grundtypen zu tun haben. Manche Grundtypen mögen nun eine gewisse Radiation erfahren haben (vgl. Abschnitt 5.3.10), andere dagegen nicht. Als Ursache für das unterschiedliche Ausmaß an Radiationen wären ökologische Bedingungen zu suchen. Auch in der heutigen Flora und Fauna können zahlreiche Beispiele für extrem unterschiedliche Ausmaße von Radiationen innerhalb von Grundtypen angeführt werden (vgl. Scherer 1993).
5.3.8 Übergangsformen an falschen stratigraphischen Positionen
In Kapitel 4 (Abschnitte 4.1.1, 4.2, 4.4.1, 4.5.2, 4.6 und 4.7.2) wurden zahlreiche Beispiele morphologisch bzw. anatomisch begründeter Zwischenformen genannt, die stratigraphisch für eine reale Übergangßtellung nicht passen; das ist sogar – wie gezeigt – der Regelfall.
|
|
||||||||
| Art/Gattung | Strat. | Taxon 1 | Strat. | Taxon 2 | Strat. | Taxon 3 | Strat. | |
|
|
||||||||
| Aglaophyton | 2 | Rhyniophytina | 0 | Moose | 6 | (4.4.1) | ||
| Horneophyton | 2 | Rhyniophytina | 0 | Moose | 6 | (4.4.1) | ||
| Nothia | 2 | Rhyniophytina | 0 | Zosterophyll. | 1 | (4.4.1) | ||
| Renalia | 3 | Rhyniophytina | 0 | Zosterophyll. | 1 | Trimerophytina | 2 | (4.4.1) |
| Hsuea | 3 | Rhyniophytina | 0 | Zosterophyll. | 1 | (4.4.1) | ||
| Taeniocrada long. | 2 | Rhyniophytina | 0 | Trimerophytina | 2 | (4.4.1) | ||
| Sawdonia spinos. | 3 | Zosterophyll. | 1 | Trimerophytina | 2 | (4.4.1) | ||
| Estinnophyton | 2 | Trimerophytina | 2 | Lycopsida | 2/0? | Cladoxylales | 3/4 | (4.2) |
| Ibyka | 5 | Iridopteridales | 4/5 | Hyenia | 3/4 | (4.5.2) | ||
| Duisbergia | 4 | Lycopsida | 2/0? | Cladoxylales | 3/4 | Pleuromeia | Trias | (4.7.2) |
| Rhymokalon | 6 | Trimerophytina | 2 | Cladoxylales | 3/4 | (4.4.1) | ||
| Eleutherophyllum | K | Lycopsida | 2/0? | Sphenopsida | 6 | |||
|
|
||||||||
Ist nun unpassende Stratigraphie ein Gegenargument gegen die Deutung als evolutionäre Übergangsform? Man kann argumentieren, daß die entsprechenden Formen auch bereits zu der Zeit existiert haben, in der sie evolutionär passend waren. Dieses Argument verliert aber mit der Anzahl der nicht passenden Fälle an Gewicht. Treten die am ehesten als Übergangsformen interpretierbaren Formen sogar, wie es scheint, systematisch an unpassenden stratigraphischen Positionen auf (vgl. Tab. 11), muß auch eine systematische Erklärung dafür gefunden werden. Daher setzt dieser Befund ein Fragezeichen an die evolutionstheoretische Interpretation. Folgt die stratigraphische Verteilung der Unterdevonpflanzen dagegen ökologischen Gründen (z. B. Besiedlung von Brachland, s. o.), ist zu erwarten, daß die Abfolge der einzelnen Gattungen nicht streng der Komplexität bzw. der Einpaßbarkeit in evolutionstheoretisch begründbare Reihen folgen.
5.3.9 Gleichzeitiges Auftreten verschiedener Varianten innerhalb eines Taxon
Der Befund des ungefähr zeitgleichen Auftretens unterschiedlich komplexer Gattungen innerhalb bestimmter Taxa (z. B. bei den Rhyniophyten, Zosterophyllophyten, Trimerophyten, Aneurophyten; vgl. Abschnitte 4.1.1 und 4.6.2) stellt eine ähnliche Anfrage an evolutionstheoretische Deutungsweisen dar wie der Befund der stratigraphisch unpassenden Mosaikformen (vgl. Abschnitt 5.3.8). Auch hier gilt, daß nicht Einzelfälle das Gewicht des Arguments ausmachen, sondern die Regelhaftigkeit dieses Befundes.
5.3.10 Mögliche Beispiele einer Grundtyp-Diversifikation
In der Rezentbiologie wurde von Scherer (1993) das Grundtypkonzept zur Diskussion gestellt. Danach werden durch Kreuzungen miteinander verbundene Arten und Gattungen zu einem Grundtyp zusammengefaßt. Es muß dabei sichergestellt sein, daß im Mischling väterliches und mütterliches Erbgut zusammenwirkt. Es wird postuliert, daß die Grundtypen ein hohes Variationspotential besaßen (polyvalentes Erbgut) und daß die heutigen Biospezies innerhalb von Grundtypen in der Regel durch Spezialisierung, d. h. genetische Verarmung entstanden. Innerhalb von Grundtypen kann es zu einer enormen, wenn auch begrenzten Vielfalt unterschiedlich spezialisierter biologischer Arten kommen.
Nun ist es naturgemäß nicht möglich, das Grundtypkonzept in der Paläontologie direkt anzuwenden. Doch kann u. U. durch den Vergleich heutiger Vielfalt innerhalb von Grundtypen auf entsprechende Verhältnisse bei fossilen Formen geschlossen werden. Vor diesem Hintergrund bietet sich die Frage an, inwieweit die devonische Vielfalt pflanzlicher Überlieferung durch Grundtyp-Diversifikation verstanden werden kann.
In Abschnitt 4.1.4 wurden die Gattungen Renalia und Täniocrada (Schweitzer 1983, 91ff.) erwähnt, die bei deutlicher Konstanz vegetativer Merkmale und des Sporangienbaus eine erhebliche Variabilität in der Sporangienstellung aufweisen. Vergleicht man diese Situation mit z.T. erheblich unterschiedlichen Blütenständen innerhalb von Grundtypen bei Angiospermen, so kann hier eine Grundtyp-Aufspaltung ohne weiteres in Betracht gezogen werden.
An unterschiedliche Varianten innerhalb von Grundtypen kann auch bei artenreichen Gattungen wie Psilophyton und Zosterophyllum gedacht werden.
Es stellt sich in dieser Sichtweise die Frage, inwieweit gewisse Unterschiede der Merkmalsausprägung auf diesem tiefen taxonomischen Niveau überhaupt signifikant für evolutionäre Tendenzen sein können. Unterschiede innerhalb eines polyvalenten Grundtyps können kaum evolutionäre Trends begründen, allenfalls einen Trend zur Spezialisierung, der aber nicht gleichzeitig ein Trend zur Höherentwicklung sein kann. Im unteren taxonomischen Bereich wäre auch zu prüfen, ob manche Mosaikformen (vgl. Abschnitt 4.1) als Hybriden gedeutet werden könnten. Für mögliche evolutionäre Tendenzen auf höherer taxonomischer Ebene dürften diese Überlegungen allerdings kaum von Belang sein.
5.3.11 Experimentelle Phase der Landpflanzenevolution?
Die genannten Schwierigkeiten in der Taxonomie devonischer Landpflanzen und beim Versuch der phylogenetischen Rekonstruktion werden oft durch eine „experimentelle Phase“ der Evolution zu erklären versucht. Die taxonomischen Schwierigkeiten seien nochmals beispielhaft anhand eines Zitats von Bierhorst (1971, 113) in Erinnerung gebracht. Er bemerkt zu den farnähnlichen devonischen Pflanzen: „At this evolutionary level of vascular plants we cannot clearly delimit orders and classes, much leß subdivisions, but the same information that disallows this adds significantly to our understanding of morphology and phylogeny.“ Familien und Ordnungen haben sich noch nicht klar herauskristallisiert; es gäbe noch keine „fixierten Typen“ (Banks 1964, 23f.) Ähnlich stellen Andrews et al. (1974, 406f.) fest: „It is now clear that the early land plants of the Lower Devonian evolved into a rather diverse assemblage by Middle Devonian times. Some of the latter ran into blind alleys or evolved into trü ferns (Filicales and Marattiales), others led to the problematical and heterogeneous Cönopterids, and others led toward the Progymnosperms, which we are just beginning to understand.“ Die Vielfalt von Strategien im Wachstum und in der Reproduktion bei den Rhyniophyten sei Ausdruck einer einmaligen evolutionären Position (Edwards & Edwards 1986, 216). Die taxonomisch nicht einordenbaren Unterdevonpflanzen seien – so Banks (1975, 411) – Zeugen der Dynamik der evolutionären Aktivität im frühen Devon-Zeitalter (ähnlich Stewart & Rothwell 1993, 127; Bateman 1991, 89). Die frühe Landpflanzenevolution ist nicht leiterförmig darstellbar, sondern eher buschig, stellen auch in jüngster Zeit Garbary et al. (1993, 254) fest (vgl. Selden & Edwards 1989). Diese Situation sei vergleichbar mit der Situation der Burgess-Shale-Fauna (Gould 1991). „Accordingly, once land plants evolved, there very quickly arose a number of evolutionary lines each with several different body plants“ (Garbary et al. 1993, 254).
Wie in den genannten Zitaten anklingt, besagt die Vorstellung von einer experimentellen Phase der frühen Landpflanzenevolution, daß die Pflanzen nach der Eroberung des neuen Lebensraumes quasi planlos in alle möglichen Richtungen evolvierten. Dabei traten in großem Maße Konvergenzen oder Parallelentwicklungen auf, die es nun schwer oder gar unmöglich machen, evolutionäre Zusammenhänge über grobe allgemeine Trends hinaus nachzuvollziehen. Dieses Konzept wirkt zunächst suggestiv. Dennoch sind dazu einige kritische Rückfragen zu stellen:
1. Bei näherem Hinsehen läuft dieser Erklärungsversuch auf die Annahme hinaus, daß beim Erobern des Festlandes zunächst die Bedingungen für das Auftreten von Konvergenzen im Vergleich zu anderen Phasen der postulierten Evolution erheblich günstiger waren. Das Schlagwort von „experimentellen Evolutionsphasen“ beinhaltet nur eine evolutionär gedeutete Beschreibung der Datenlage und noch keine Erklärung. Auch die folgende Erklärung von Banks ist nur eine Beschreibung des Phänomens: „External morphology, mode of reproduction and anatomical structure were evolving at different rates and in different directions“ (Banks 1964, 19). Um über eine bloße Beschreibung hinauszukommen, muß verständlich gemacht werden, weshalb in dieser Phase das gehäufte Auftreten von Konvergenzen begünstigt war. Es sollte eine Erklärung dafür geben, weshalb unterschiedliche Pflanzenorgane unterschiedlich schnell evolvierten, und weshalb in verschiedenen Gruppen dieselben Organe verschieden schnell evolvierten (Beispiele bei Stein et al. 1984). Es läge auch das Argument nahe, daß eine experimentelle Phase im Gegensatz zum vorliegenden Befund nur wenig Konvergenzen hervorbringen sollte (nämlich aufgrund von „evolutionären Versuchen“ in verschiedene Richtungen). Eine Antwort könnte unter Berücksichtung konstruktionsmorphologischer Gesichtspunkte gefunden werden. Vielleicht ermöglichten die frühen Landpflanzenkonstruktionen nur bestimmte Entwicklungsrichtungen.
2. Die Tragfähigkeit des Arguments muß sich an konkreten Gattungen bewähren. Die Vorstellung einer experimentellen Phase macht das relativ späte Auftreten von Mosaikformen (vgl. Tab. 11) nicht verständlich; man denke etwa an Renalia (Abschnitt 4.1.1), Rhymokalon (Abschnitt 4.4.1) oder Duisbergia (Abschnitt 4.7.2). Ebensowenig wird das frühe Auftreten abgeleiteter Formen erklärt (Zosterophyllum myretonianum, Yarravia, Psilophyton crenulatum, Tetraxylopteris, Barinophyton, Estinnophyton).
3. Die Befunde, die die Vorstellung einer experimentellen Phase begünstigen, treten nicht nur in der Phase der Eroberung des Landes auf, sondern auch später. So weisen die Samenanlagen-Typen des Unterkarbons eine recht große Vielfalt auf; sie sind aber köxistent; eine evolutionäre Reihe kann nicht gebildet werden (DiMichele 1989, 7, 8, 10). Eine vergleichbare Situation trifft auf die Vielfalt der Blattformen der Sphenophyllales zu (Meyen 1987, 88).
Auch in der Anfangsphase der Landeroberung, also beim eigentlichen Schritt ans Land noch vor der Radiation der Flora auf dem Land, wird angesichts der Vielfalt morphologischer und physiologischer Strategien, die nicht in unser Konzept rezenter Landpflanzen (Tracheophyten und Bryophyten) fallen, das Konzept der experimentellen Evolution bemüht (Edwards & Fanning 1985, 154f., ähnlich Edwards 1986, 201).
5.3.12 Fehlen von Übergangsformen
Einerseits liegen mit zahlreichen Mosaikformen quasi zuviele „Übergangsformen“ vor, andererseits sind andere Taxa recht isoliert, und über evolutionäre Zusammenhänge kann nur spekuliert werden. Das gilt für die Sphenopsiden-Ordnungen, insbesondere für die Pseudoborniales (Abschnitt 4.5.5), für die Taxa incertä sedis (Abschnitte 4.3, 4.6.6, 4.7.2), aber auch in unterschiedlicher Deutlichkeit für zahlreiche andere Pflanzengruppen (wie in Kapitel 4 im einzelnen diskutiert).
5.3.13 Evolutionäre Stasis
In Überblickskapitel 3 wurde bereits erwähnt, daß eine Reihe von Gattungen über längere Zeiten stabil bleibt (Abb. 8). So weist Cooksonia pertoni vom Wenlock (Obersilur) bis Unterdevon abgesehen von der Sporenmorphologie keine Veränderungen auf (Fanning et al. 1988, 23); Sawdonia persistiert nahezu unverändert vom Siegen bis zum Frasne (Gensel & Andrews 1987, 484); Huvenia ist fast konstant vom höheren Siegen bis zum höheren Ems (Schultka 1991, 174), ebenso auch Leclercqia vom Ems bis zum Givet (Banks 1985, 198). Merkmale, die viele Arten der heutigen Gattung Lycopodium auszeichnen, sind seit Baragwanathia und anderen damals lebenden Lycopsiden (Obersilur oder Unterdevon) bemerkenwert unverändert: kriechend-aufsteigender Habitus; dichotome und / oder anisotome Verzweigung, zahlreiche spiralig angeordnete Mikrophylle, Protostele, Sporen in großen Sporangien, Sporangien in adaxialer Stellung zu den Blättern. Was hat zu dieser Konstanz beigetragen in einer Zeit, die von vielen Autoren als „experimentelle Phase“ der Landpflanzenevolution angesehen wird? (vgl. Abschnitt 5.3.11) Diese Frage stellt sich besonders angesichts der Tatsache, daß die stabilen Arten in denselben Biotopen gelebt haben wie diejenigen, die als Zeugen gewaltiger evolutionärer Übergänge betrachtet werden.
5.3.14 Sind die Deutungsprobleme im Rahmen der Evolutionslehre auf die Lückenhaftigkeit der Fossilüberlieferung zurückzuführen?
Solange die Fossilüberlieferung eher dürftig ist, kann zur Erklärung des Fehlens von Zwischenformen auf das alte Argument der Lückenhaftigkeit der Fossilüberlieferung zurückgegriffen werden. Es wird aber mit zunehmender Zahl von Fossilfunden unglaubwürdig. Da die Artenzahl unter den Devonpflanzen tatsächlich relativ dürftig ist, kann man dieses Argument vorerst gelten lassen. Die weitere Forschung wird Klarheit bringen müssen.
In den zahlreichen Fällen aber, wo die überlieferte Formenvielfalt häufige Heterobathmien und Konvergenzen belegt, hilft der Verweis auf die Lückenhaftigkeit der Fossilüberlieferung nicht weiter – im Gegenteil: Es wurde bereits darauf hingewiesen, daß die bereits festzustellende Zahl an Konvergenzen eher noch zunehmen dürfte, wenn noch weitere Taxa bekannt oder die bereits bekannten Taxa in vollständigerer Erhaltung überliefert werden. Es mag durch weitere Funde gelingen, eher eine Entscheidung darüber zu treffen, welche Merkmale als konvergent zu betrachten sind. Nach dem derzeitigen Kenntnißtand kann diese Entscheidung oftmals nicht getroffen werden (vgl. Stein et al. 1984).
5.3.15 Zeitraum der Fossilüberlieferung und Entstehungszeitraum
Verschiedentlich wurde die Vermutung geäußert, daß die Orte der Überlieferung der devonischen Landpflanzen nicht mit den Gebieten ihrer evolutiven Entstehung gleichzusetzen seien. „Time of evolution and time of appearance in the fossil record must be kept conceptually distinct in any consideration of land-plant evolution“ (Gray & Boucot 1977, 148; Hervorhebung im Original). Zu dieser Problematik seien im folgenden einige Übelegungen angestellt.
Ökologische Aspekte. Die Abfolge der Fossilüberlieferung kann grundsätzlich außer evolutionären auch ökologische Gründe haben (vgl. Abschnitt 5.2). Beispielsweise können sich homospore Pflanzen schneller ausbreiten als heterospore oder samentragende. Holzige Pflanzen müssen mehr Energie für nicht-photosynthetisierende Gewebe aufbringen als krautige Pflanzen (vgl. Raven 1984). In Prionierbiotopen, in denen es auf schnelle Reproduktionsreife ankommt, sind krautige, kleinwüchsige und schnellreifende Pflanzen wie Cooksonia (s. Abb. 1) daher bevorzugt. Unter minimalem Aufwand an Energie und bei Verzicht auf sekundäre Gewebe ist eine Sporenproduktion am schnellsten möglich (Edwards & Fanning 1985, 157). Blattlosigkeit könnte in einem Zusammenhang mit UV-Belastung und in einem Zusammenhang mit klimatischen Störungen stehen. Eine ökologisch bedingte Sukzession könnte also das zeitlich gestaffelte Auftreten von Merkmalen und Pflanzengruppen u. U. durchaus erklären. Die in den vorigen Abschnitten angeführten Deutungsschwierigkeiten im Rahmen der Evolutionslehre könnten in einem katastrophischen Kurzzeitrahmen evtl. leichter zu deuten sein, nämlich die Gleichzeitigkeit des Auftretens verschiedener Organisationsformen oder die Existenz „stratigraphischer Ausreißer“.
Geologische Aspekte. Das Spektrum der Überlieferung von Pflanzenresten kann auch geologische Gründe haben. Pflanzen in kontinentalen Sedimenten könnten eher durch nachfolgende Erosion zerstört worden sein als Küstenvegetation (Axelrod 1959). Dieser Autor hält es für möglich, daß die Phasenabfolgen der devonischen Floren nichts anderes repräsentieren könnten als Wanderungen von ökologischen Einheiten von höheren in tiefere Lagen und nicht auf evolutionäre Änderungen als solche zurückgehen, sondern nur eine Ablösung verschiedener ökologischer Einheiten belegen (Axelrod 1959, 270). Er weist in diesem Zusammenhang darauf hin, daß die großen Mengen nicht metamorphisierter frühpaläozoischer und präkambrischer Sedimente marinen und nicht kontinentalen Ursprungs sind, so daß dort kaum Landpflanzenreste zu erwarten seien (S. 272). Ähnlich urteilt Leclercq (1956, 110), daß auf der Grundlage der Makrofossil-Flora ökologische Lebensgemeinschaften irrtümlich als evolutionäre Stadien interpretiert werden könnten. Axelrod geht davon aus, daß die Pflanzenwelt auf dem Niveau von Devonpflanzen 150-200 Millionen Jahre in höheren Regionen wohl existierte, aber aufgrund der Sedimentationsbedingungen nicht konserviert wurde und unentdeckt blieb. Kontinentale Formationen sind generell ungeeignet für eine Erhaltung von Pflanzenabdrücken, so Leclercq (1956, 112). Diesen Zusammenhang diskutiert auch Edwards (1970, 112). Bestritten wird diese Erklärung durch Chaloner (1967). Weder Mikro- noch Makrofossilien würden eine vorsilurische Landpflanzenevolution belegen. Die Migrationshypothese von Axelrod sei zwar nicht von vornherein unplausibel, jedoch würde die Beobachtung der Größenänderungen der fossilisierten Sporen diese These sehr unplausibel machen (Chaloner 1967, 87, 89f.). Chaloner verweist auf Regelhaftigkeiten der Sporenüberlieferung. In einem Langzeitrahmen (also einer über Millionen Jahre sich erstreckenden frühen Landpflanzenentwicklung) sollte angesichts der leichten Verbreitung keine Sporensukzession (in marinen Sedimente) erwartet werden, wenn verschiedene Landpflanzentypen bereits gleichzeitig existieren (aber nicht als Makrofossilien dokumentiert sind).
In einem Kurzzeitrahmen könnte die Deutung Axelrods jedoch wieder an Plausibilität gewinnen. Die Sukzession in der Mikrofossilüberlieferung könnte dann auf eine sukzessive Land-(wieder-)besiedlung zurückzuführen sein. Zu beachten ist auch, daß das inzwischen bekannte Ausmaß der Mikrofossilüberlieferung die Existenz von silurischen und evtl. sogar ordovizischen Makrofossilien doch plausibel erscheinen läßt.
Edwards & Fanning (1985, 149) greifen in jüngerer Zeit Axelrods Überlegung wieder auf: Für die frühe devonische Flora bleibe offen, ob sie auf sukzessive Migrationen zurückgehe, während ihre Evolution an anderen Orten erfolgte. Die Autoren halten die Mikrofossilien für die entscheidenden Belege der Evolution, allerdings blieben Zuordnungen zu Makrofossilien im Bereich der Vermutungen.
Der neue Fund von Pinnatiramosus (vgl. Kapitel 3, 1.) könnte diese Diskussion neu beleben, denn diese Gefäßpflanze aus dem unteren Silur paßt nicht ins etablierte evolutionstheoretische Szenario. Cai et al. (1996) plädieren denn auch dafür, das Bild von der frühen Landpflanzenevolution zu revidieren. Die weitere Forschung verspricht spannend zu werden.
Auch Gray & Boucot (1971, 921) halten es für möglich, „that the known progressive increase in the frequency and taxonomic diversity of trilete spores that occurs from the Early Silurian to the Late Silurian-Early Devonian may be largely a function of biofacies correlated with geologic environment rather than a function of organic evolution as now concluded.“ Sie sprechen von einem „environmental artifact“. Die silurischen Sporen könnten von Gefäßpflanzen stammen. „. . . it may yet be found that Early Paleozoic vascular plants show a trend similar to the evolutionary history of many animal groups, which exhibit far greater diversity and morphological richneß in their early rather than their later history“ (Gray & Boucot 1971, 921). Sporen, die ohne weiteres zu Landpflanzen gehören könnten, finden sich ab dem späten Ordovizium weltweit (Gray et al. 1974, 261; Gray & Shear 1992).
Schließlich sei noch ein interessantes Einzelbeispiel genannt: Die Abfolge der taxonomischen Diversität in Gotland und der Reichtum vordevonischer Sporen ist eher mit der Umgebung korreliert als mit dem geologischen Alter. Wäre Evolution der allein maßgebliche Faktor, wäre eine strenge Korrelation zwischen der Sporenmenge und ihrer Vielfalt und der Alter der Sedimente zu erwarten. Die jüngeren Sedimente sollten die größte Sporenvielfalt und zahlreiche Sporen aufweisen, doch das tun sie gerade nicht. Für keine silurische Sektion ist eine beständige durchschnittliche Zunahme an Zahl und morphologischer Diversität von Sporen von der Basis bis zur Spitze festzustellen. Die Unterschiede in der taxonomischen Diversität der Sporen scheint nur mit den Umgebungen der Ablagerungsgebiete zusammenzuhängen, nicht jedoch mit dem Evolutionßtadium der Mutterpflanzen (Gray & Boucot 1977, 171).
Die Hinweise auf die Existenz von Landpflanzen deutlich vor ihrer fossilen Überlieferung scheinen sich mehr und mehr zu verdichten. So berichten neuerdings Remy et al. (1994) von einer vesiculär-arbuskulären Mykorrhiza (die heutigen Mykorrhizen gleicht) bei der unterdevonischen Aglaophyton major (vgl. Abschnitt 4.1; Abb. 10). Die Entstehung dieser Symbiose muß in einer entsprechend frühen Zeit postuliert werden – ein indirekter Hinweis auf eine frühe Existenz von Land(gefäß-) pflanzen. Ebenso weisen Spurenfossilen von Gliedertieren aus dem Ordovizium auf frühe Landbesiedlung hin (Johnson et al. 1994; Gray & Shear 1992). Die Diskussion um den Zeitraum der Landpflanzenentstehung kann nicht als abgeschlossen angesehen werden.
5.3.16 Wie gut bewährt sich die Telomtheorie?
Abschließend sollen noch einige zusammenfassende Bemerkungen zur Telomtheorie gemacht werden. Diese sehr einflußreiche übergreifende Theorie zur Erklärung evolutionärer Veränderungen in der frühen Landpflanzenevolution wurde bei den Einzelbetrachtungen bereits gestreift. Nach der von Zimmermann begründeten Telomtheorie (vgl. Zimmermann 1965) soll durch wenige relativ einfache „Elementarprozesse“ die gestaltliche Vielfalt von Landpflanzen ausgehend von einfachen, nackten Gabelsprossen verständlich gemacht werden (Abb. 52). Im einzelnen handelt es sich um Übergipfelung, Planation, Verwachsung, Inkurvation (Einkrümmung) und Reduktion. Die Verwachsung kann flächig oder dreidimensional erfolgen.
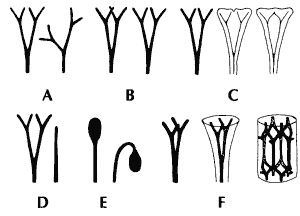
Abb. 52: „Elementarprozesse“ nach der Telomtheorie. A Übergipfel, B Planation, C Verwachsung im Blatt, D Reduktion, E Einkrümmung, F Verwachsung in der Achse (Nach Zimmermann 1965)
Inwieweit hilft nun die Telomtheorie zum Verständnis der morphologischen Ausprägungen bei den Devonpflanzen? Zweifellos passen viele Pflanzengestalten oder einzelne Organe zu den Postulaten der Telomtheorie. Man kann hier an verschiedene Grade der Übergipfelung, der Planation, Verwachsung und der Reduktion von Zweigen sowie an die Einkrümmung von Sporangienträgern denken. An einigen konkreten Beispielen zeigt sich jedoch, daß die Telomtheorie kaum eine allgemeingültige Erklärung für postulierte evolutionäre Abfolgen bietet und daß Befunde vorliegen, die sich gegen eine telomtheoretische Deutung sperren. Im folgenden sollen einige Beispiele Revü passieren, die bereits in Kapitel 4 genannt worden waren.
- Rhynia gwynne-vaughanii (Abb. 1) zeigt ein relativ kompliziertes Verzweigungssystem mit Adventivzweigen; Edwards (1980, 185) hält es für möglich, daß entgegen der Telomtheorie monopodiale Verzweigung mindestens in einigen Fällen von solchen Adventivzweigen ihren Ausgang nahm, nicht durch Übergipfelung.
In diesem Sinne äußert sich auch Remy (1986, 1) kritisch: Die Vielfalt in Form und Ausbildung von Organen bereits im Unterdevon lasse sich nur selten befriedigend mit der Telomtheorie erklären. Es komme darauf an, das Geschehen am Sproßscheitel zu rekonstruieren; hier gebe es eine Vielzahl von Abwandlungsmöglichkeiten. Daher müßten sog. „evolutionäre Reihen“ kritisch bewertet werden (Remy 1986, 5). Das heißt also: Abfolgen, bei denen einfach Adultstadien aneinandergereiht werden, setzen methodisch verkehrt an.
- Das geteilte bis gelappte Sporangium von Horneophyton (Abb. 11) könnte Vorläufermodell von Synangien sein (statt Zusammenfassung einzelner Sporangien nach der Telomtheorie; Eggert 1974, 405).
- Zosterophyllophyten: Das sehr frühe Auftreten stengelständiger Sporangien paßt nicht zur Telomtheorie, nach deren Vorstellung diese Position durch starke Reduktion von Telomen zu erklären wäre, also einen abgeleiteten Zustand darstellen würde.
- Die Ableitung der Lycopsiden-Mikrophylle ist zumindest umstritten. Auch hier spricht das frühe Auftreten echter (vaskularisierter) Mikrophylle gegen die telomtheoretische Vorstellung der Entstehung. In Abschnitt 4.2 wurden Gründe genannt, die für eine Ableitung von Emergenzen (Stacheln oder dergleichen) sprechen.
- Die Entstehungsweise der blattständigen Sporangien ist ebenfalls umstritten. Neben der telomtheoretischen Deutung durch Reduktion sporangientragender Zweige wird auch eine allmähliche Verschiebung der stengelständigen Sporangien auf das Mikrophyll diskutiert.
- Die Blätter der Paläophyllales (Abb. 23) sind vollständig flächig verwachsen. Dabei handelt es sich um einen Endzustand nach der Telomtheorie. Auch hier tritt diese Ausprägung stratigraphisch viel zu früh auf, wenn die Telomtheorie als Maßstab genommen wird.
- Calamophyton und Hyenia (vgl. Abschnitt 4.5.1) wurden u. a. wegen der Inkurvation der Sporangienträger als Vorläufer der Sphenopsiden diskutiert, doch hat sich diese Deutung als wenig tragfähig erwiesen. Ob damit die Inkurvation von Sporangien als Vorstufe zu den sphenopsidischen Sporangientischchen gewertet werden kann, bleibe dahingestellt. Die Hyeniales als vormalig wichtige Belege sind dafür jedenfalls hinfällig geworden.
