
Evolutionstheorie(n) in der Krise?
30.11.22 Ist Lamarck zurück?
Die Vererbung erworbener Eigenschaften ist bei Korallen doch möglich
Nach gängiger Auffassung kann eine neue Mutation nur dann zu evolutionären Veränderungen beitragen, wenn sie in Zellen des Fortpflanzungssystems auftritt, d. h. in einer Ei- oder Samenzelle. Mutationen in allen anderen Körperzellen gelten als evolutionär irrelevant, da sie nicht an die Nachkommen vererbt werden. Diese Sichtweise scheint nicht durchweg zu stimmen. Zumindest bei Korallen ist die Vererbung erworbener Merkmale ein Mechanismus für Veränderungen nun tatsächlich nachgewiesen. Dies bestätigt teilweise die Ansichten des Naturforschers Jean Baptiste de Lamarck im 19. Jahrhundert, welche man ca. ein Jahrhundert ignoriert hatte.
Im Jahr 1809 stellte Jean-Baptiste de Lamarck eine erste ausgearbeitete Evolutionshypothese auf, die sich weitgehend auf die Vererbung erworbener Merkmale stützte. Er argumentierte, dass sich Organismen im Laufe des Lebens an bestimmte ökologische Herausforderungen anpassen würden, und dass diese Anpassungen an die Nachkommen vererbt werden könnten.
1892 veröffentlichte August Weismann (1834–1914) die Hypothese, dass somatische Mutationen, die während des Lebens eines Tieres in den Körperzellen erworben werden, evolutionär irrelevant sind, weil sie die angenommene Barriere (die so genannte Weismann-Schranke) zwischen Soma (Körperzellen) und Keimbahn (Keimzellen) nicht überwinden und somit nicht zur genetischen Variation der nächsten Generation beitragen können. Er widersprach damit der Möglichkeit der Lamarck‘schen Evolution: Von den Eltern erworbene Merkmale könnten niemals zu an die Nachkommen vererbt werden.
Alle nachfolgenden Evolutionstheorien stützten sich stark auf die Weismann-Schranke, wonach weder die Körperzellen eines Organismus noch die Umwelt das genetische Erbe beeinflussen können. Anpassungen sind insofern zufällig, als Variation (infolge von Mutationen des Erbgutes) zufällig ist. Die zweite treibende „Kraft“ der Darwin’sche Evolution ist die natürliche Auslese, wonach die am besten Angepassten in der Regel die meisten Nachkommen haben. Auf diese Weise soll Evolution erfolgen.
Auch wenn die Geschlechtszellen vollständig von den Körperzellen getrennt sein sollten, könnte es dennoch molekular-genetische Mechanismen geben, durch die die Geschlechtszellen über die Umwelt informiert werden und die Nachkommen dadurch mit möglichen Anpassungen vorbereitet werden. Dass dies tatsächlich der Fall ist, wurde inzwischen in der Literatur ausführlich dokumentiert (Lacal & Ventura 2018, Miryeganeh & Saze 2020, Triantaphyllopoulos et al. 2016). Man spricht von epigenetischer Vererbung. Epigenetisch vererbbare Merkmale sind jedoch oft nicht mit Mutationen in der DNA verbunden, sondern betreffen in der Regel die Regulation der DNA. Ob sie stabil über viele Generationen vererbbar sind, ist bisher weitgehend ungeklärt.
Bei sich sexuell fortpflanzenden Organismen verschwinden vorteilhafte Mutationen der DNA in der Regel durch zufällige genetische Drift und werden fast nie in der Population fixiert. Nur wenn die Selektion sehr stark ist, d. h. wenn die Mutation einen enormen Reproduktionsvorteil bringt, kann sich eine vorteilhafte Mutation häufiger in der gesamten Population durchsetzen. Es wäre daher von großem Wert, wenn adaptive (vorteilhaft angepasste) Veränderungen direkt, also ohne Beteiligung von Geschlechtszellen vererbt werden könnten. Dies geschieht bei der ungeschlechtlichen Fortpflanzung wie z. B. bei Ablegern von Pflanzen, wenn die Mutationen zuvor in den für die Ableger-Bildung zuständigen Zellen aufgetreten sind. Wäre dies auch bei der sexuellen Fortpflanzung möglich, würde das dem Lamarck‘schen Konzept der genetischen Anpassung entsprechen. Somatisch erworbene Merkmale müssten auf irgendeinem Wege auch auf die Geschlechtszellen übertragen werden, damit sie dauerhaft vererbbar bleiben.
Entdeckung einer Lamarck’schen Vererbung
Die Entdeckung eines internationalen Wissenschaftlerteams unter der Leitung von Biologen der Penn State University offenbart nun tatsächlich einen neuartigen Mechanismus zur Anpassung an sich rasch verändernde Umweltbedingungen, der dem Lamarck’schen Mechanismus sehr ähnlich ist (Kuntz et al. 2022). Das Forschungsteam analysierte die DNA-Unterschiede in einer großen Kolonie von Elchgeweih-Korallen (Acropora palmata) mit Nachkommen eines einzigen Elternteils (also mit eingeschlechtlicher Fortpflanzung) sowie in DNA-Proben von fünf benachbarten Kolonien.
Wie die meisten Korallen können sich auch die Elchgeweih-Korallen sowohl ungeschlechtlich (durch Knospung) als auch geschlechtlich (durch Produktion von Ei- und Samenzellen) fortpflanzen. Bei den hier untersuchten Elchhornkorallen, die ihre Ei- und Samenzellen beim Laichen ins Wasser abgeben, werden die Eier einer Korallenkolonie in der Regel durch Spermien einer benachbarten Kolonie befruchtet. Das Forscherteam fand jedoch heraus, dass sich einige Eier der Elchhornkoralle auch ohne Befruchtung zu lebensfähigen Nachkommen entwickeln können.
Die Ergebnisse zeigten, dass die separaten Korallenkolonien (also die o. g. benachbarten Kolonien) zu derselben ursprünglichen Koralle gehört haben. Dies bedeutet, dass es sich um Klone handelt, die von einer einzigen Originalkolonie abstammen, die sich durch ungeschlechtliche Fortpflanzung (Knospung) vermehrt hat. Jede genetische Variation, die bei den heutigen Korallen in diesen Kolonien gefunden wurde, muss also das Ergebnis einer somatischen Mutation gewesen sein.
Das Team fand insgesamt 268 somatische Mutationen in den Proben, wobei jede Korallenprobe zwischen 2 und 149 somatische Mutationen aufwies. Das Team untersuchte außerdem die Ein-Eltern-Nachkommen der Elchkorallen-Elternkolonie und stellte fest, dass 50 Prozent der somatischen Mutationen vererbt worden waren (Kuntz et al. 2022).
Der genaue Mechanismus, wie die somatischen Mutationen ihren Weg in die Keimzellen der Korallen finden, ist noch unbekannt, aber die Forscher vermuten, dass die Trennung zwischen Körper- und Keimzellen in Korallen unvollständig ist und einige Körperzellen die Fähigkeit behalten, Keimzellen zu bilden, wodurch somatische Mutationen ihren Weg in die Nachkommen finden können (Penn State 2022). Ein ähnlicher Mechanismus, der von John A. Davison unter dem Begriff „semi-meiotische Hypothese“ ausführlich erörtert wurde, könnte ein relevanter nicht-darwinistischer Mechanismus für Anpassungen und Artbildung sein (Davison 1984).
Einer der Autoren kommentierte: „Da Korallen als Kolonien genetisch identischer Polypen wachsen, können somatische Mutationen, die in einem Korallenpolypen entstehen, der Umwelt ausgesetzt und auf ihren Nutzen hin untersucht werden, ohne notwendigerweise die gesamte Kolonie zu beeinträchtigen. Daher können Zellen mit potenziell schädlichen Mutationen sterben und Zellen mit potenziell vorteilhaften Mutationen können florieren und sich ausbreiten, während die Korallenkolonie weiterwächst. Wenn diese Mutationen dann an die Nachkommen weitergegeben werden können – wie wir jetzt gezeigt haben – bedeutet dies, dass die Korallen ein zusätzliches Werkzeug haben, das ihre Anpassung an den Klimawandel beschleunigen könnte“ (Penn State 2022).
Spätestens hier bricht die Weismann-Schranke zusammen: Die Vererbung von somatischen Mutationen ist Lamarck’sche Evolution, und keine neodarwinistische Evolution.
Eine hässliche Tatsache oder schöne Wissenschaft?
Es war Thomas Henry Huxley, ein überzeugter Verfechter der Darwin‘schen Thesen, der vor über 100 Jahren schrieb: „Die große Tragödie der Wissenschaft – die Erschlagung einer schönen Hypothese durch eine hässliche Tatsache.“ Man sollte sich darüber im Klaren sein, dass Fakten oder Beobachtungen von Naturprozessen niemals hässlich sind. Naturwissenschaftliche Befunde, die nicht in das herrschende Paradigma passen, sind ein wichtiger Faktor, der wissenschaftliche Erkenntnisse vorantreibt. Diese neue Erkenntnis bei den Korallen ist eine solche „hässliche Tatsache“: Sie zeigt uns, dass man der Wissenschaft immer damit rechnen muss, dass bisher gesicherte Theorien nicht immer von Dauer sind, sondern nur vorläufige Gültigkeit haben. Ob Lamarck´sche Mechanismen bei Anpassungsprozessen in anderen Organismen eine große Rolle spielen oder spielten, wird dank dieser „hässlichen Tatsache“ nun Gegenstand von Forschung und Diskussion sein.
Quellen
Davison JA (1984) Semi-meiosis as an evolutionary mechanism. J. theor. Biol. 111, 725–735. https://www.sciencedirect.com/science/article/abs/pii/S0022519384802647
Kuntz KLV et al. (2022) Inheritance of somatic mutations by animal offspring. Sci. Adv. 8:35; doi:10.1126/sciadv.abn0707
Lacal I & Ventura R (2018) Epigenetic Inheritance: Concepts, Mechanisms and Perspectives. Front. Mol. Neurosci. 28, https://www.frontiersin.org/articles/10.3389/fnmol.2018.00292/full
Miryeganeh M & Saze H (2020) Epigenetic inheritance and plant evolution. Popul. Ecol. 62, 17–27. https://esj-journals.onlinelibrary.wiley.com/doi/10.1002/1438-390X.12018
Penn State (2022) Corals pass mutations acquired during their lifetimes to offspring. Science Daily, https://www.sciencedaily.com/releases/2022/08/220831152728.htm
Triantaphyllopoulos KA, Ikonomopoulos J & Bannister AJ (2016) Epigenetics and inheritance of phenotype variation in livestock. Epigenetics & Chromatin 9:31. https://epigeneticsandchromatin.biomedcentral.com/articles/10.1186/s13072-016-0081-5
Autor dieser News: Peter Borger
© 2022, http://www.genesisnet.info/schoepfung_evolution/n309.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
01.08.22 Der unterschätzte Vater der Genetik: zu Gregor Mendels 200. Geburtstag
Gregor Mendel gilt als Vater der Vererbungslehre (Genetik). Dieses Jahr jährt sich sein Geburtstag zum 200. Mal. Wenig bekannt ist, dass Mendels Ideen zur Artbildung sehr gut in ein Schöpfungskonzept passen und sich in aktuellen Studien als fruchtbar erweisen.
Kann ein 160 Jahre altes Werk eines Naturforschers heute noch Aktualität besitzen? Vielen wird zu dieser Frage bestimmt Charles Darwin einfallen. Dessen Hauptwerk „Über die Entstehung der Arten“ wird bis heute als großer Durchbruch in der Biologie gewertet. Es gibt aber ein Werk eines Zeitgenossen von Darwin, dessen Relevanz für das Verständnis des Artenwandels heute kaum jemandem bewusst ist. Die Rede ist von Gregor Mendel, der auch als Vater der Genetik bezeichnet wird. Im Jahr 2022 jährt sich sein Geburtstag zum 200. Mal – Anlass für viele Beiträge in der Wissenschaftsliteratur und in den Wissenschaftsnachrichten. Hier und da wird dabei auch das Verhältnis von Mendel und Darwin thematisiert.
Darwin und Mendel. Die auf Darwin zurückgehende Erklärung für die Entstehung von Arten unterscheidet sich grundlegend von Mendels Ansatz. Im Gefolge Darwins werden geringfügige Veränderungen und Auslese als Hauptfaktoren angesehen. Heute werden als Quelle der Veränderungen ungerichtete Mutationen (spontane Änderungen des Erbguts) angesehen. Mutationen können sich – wenn überhaupt – erst im Laufe vieler Generationen in der Population durchsetzen, und für nennenswerte Veränderungen werden viele aufeinanderfolgende Mutationen benötigt. Daher müssen große Zeiträume für die Bildung neuer Arten veranschlagt werden. Mendels Konzept dagegen beruht auf präexistenten (bereits vorliegenden) genetischen Programmen. Diese ermöglichen Änderungen innerhalb weniger Generationen, da Unterschiede zwischen den Arten nicht in langwierigen Prozessen aufgebaut werden müssen, sondern von Beginn an bereits angelegt sind (Crompton 2019). Diese Veränderungen bewegen sich dabei aber nur innerhalb abgegrenzter Formenkreise, die man als genetische Familien bezeichnen kann. Diese wiederum entsprechen ungefähr den „Grundtypen“, die im Rahmen der biblischen Schöpfungslehre auf geschaffene Arten zurückgeführt werden.
Da nun beide Ansätze sehr verschieden sind, klingt es überraschend, wenn das Online-Wissenschaftsmagazin Scinexx behauptet, Mendel habe in seinen Schriften „durchaus ähnliche Vorstellungen wie Darwin und seine Mitstreiter“ vertreten (Podbregar 2022). „Mendel beschrieb ein darwinistisches Szenario einer natürlichen Selektion und eines ‚Kampfs ums Dasein‘“, wird Daniel Fairbanks von der Utah Valley University zitiert (Fairbanks 2020). Und: „Wenn man die existierenden Belege als Ganzes betrachtet, dann ergibt sich von Gregor Mendel das Bild eines akribischen Forschers, der die Grundsätze der darwinistischen Evolution akzeptierte.“ Fairbanks (2020) stellt aber auch fest, dass Mendels Versuche die Vorstellungen von Darwin über Vererbung widerlegten und zudem bewiesen, dass die Vererbung auf der freien Kombination von unteilbaren Einheiten beruhte.
Kann man Mendel mit Darwin versöhnen? Kann man die widersprüchlichen Einschätzungen über Mendel und Darwin zusammenbringen? Es ist möglich, wenn man bedenkt, dass in Mendels Ansatz die Darwin’schen Mechanismen nur eine untergeordnete Rolle spielen. Darwin wies nach, dass es eine phänotypische Variation der Arten gibt. Dem stimmte Mendel zu. Für Darwin war natürliche Auslese die Ursache für Veränderungen und Artbildung, er wusste aber nicht, wie die genetische Vielfalt entstand. Mendel stimmte zwar ebenfalls zu, dass es natürliche Auslese gibt, doch er sah sie nicht als Ursache für Artbildung. Der entscheidende Unterschied zwischen Darwin und Mendel betrifft die Quelle der Variation, die benötigt wird, bevor man überhaupt etwas selektieren kann. Mendel zeigte, dass die Quelle der Variation die Rekombination von bereits bestehenden phänotypischen Programmen war. Somit besteht nach Mendels Modell keine Notwendigkeit, neue Merkmale (durch Mutationen) zu schaffen, damit neue Arten entstehen können. Arten entstehen durch Neukombination schon vorhandener Merkmale. Was Mendel bereits über diese latenten Merkmale wusste und was man mit heutigem biologischem Wissen dazu sagen kann, hat Crompton (2019) ausführlich beschrieben. Leider wurde Mendel in diesem entscheidenden Punkt von der wissenschaftlichen Gemeinschaft übersehen. Stattdessen schlug man einen anderen Weg ein und sah Mutationen als Quelle der Variation an.
Ein weiterer grundlegender Unterschied zwischen Darwin und Mendel ist, dass nach Mendel die Variation, die durch die latenten Merkmale ausgeprägt kann, begrenzt ist und nicht ausreicht, um den Ursprung allen Lebens zu erklären. Erklärt werden kann nur die Variation, die in genetischen Familien beobachtet wird.
Der empirische Test. Wer hat nun aber recht, Mendel oder Darwin? Geht man von Beobachtungen aus, die in den letzten etwa 50 Jahren bei Langzeitstudien im Freiland gemacht wurden, so ist das Modell der Mendel’schen Artbildung klar im Vorteil. Denn es mehren sich die Beispiele für sehr schnelle Merkmalsänderungen und sogar Artbildungen, die nur möglich sind, wenn ein genetisches Potenzial an Veränderungsmöglichkeiten bereits vorhanden ist. „Schnell“ heißt hier: Innerhalb Jahrzehnten oder Jahrhunderten und sogar innerhalb weniger Generationen. Nur auf Basis präexistenter Programme sind derart schnelle, vorteilhafte Veränderungen möglich. Ein sehr lehrreiches Beispiel sind die Eidechsen der Gattung Anolis, die auf den karibischen Inseln vorkommen. Von diesen Eidechsen gibt es hunderte Arten. Man kann sie in wenige sogenannte Ökomorphe einteilen; das sind Formen, die verschiedene Lebensräume bevorzugen und entsprechende Unterschiede im Körperbau zeigen. Durch genetische Studien konnte gezeigt werden, dass die Ökomorphen auf den Inseln vielfach unabhängig entstanden sind. Außerdem haben Experimente zur Wiederbesiedlung kleiner Inseln durch Anolis-Eidechsen gezeigt, dass die Entstehung verschiedener Ökomorphe innerhalb weniger Generationen erfolgen kann. Insgesamt kann man diese Befunde am besten so erklären: Die AnolisEidechsen weisen eine präexistente und zum Teil verborgene genetische Vielfalt auf, die unter bestimmten Lebensbedingungen ziemlich schnell abgerufen werden kann. Das entspricht dem Ansatz von Mendel und widerspricht Darwins Theorie.
Rasche und wiederholte Radiationen dieser Art wurden in den letzten Jahren vermehrt beobachtet oder indirekt nachgewiesen und erweisen Mendels Ansatz als fruchtbar und dem Darwin’schen Ansatz überlegen. Aus der Sicht der Schöpfungslehre liegt es nahe, die präexistente Vielfalt der Ausgangsgruppen solcher Radiationen als geschaffenes Variationspotenzial zu postulieren. Die geschaffenen Arten waren demnach mit Variationsprogrammen ausgestattet, die vielfältige Anpassungen ermöglicht haben.
Darwin ohne Mendel. Mendel hat Darwins Arbeiten gekannt, wie Randnotizen in Mendels Handschrift belegen. Aber umgekehrt war das nicht der Fall. „Offenbar hat Darwin nie ein Exemplar von Mendels Fachartikel zu den Pflanzenhybriden erhalten und auch keine Sekundärliteratur dazu gelesen“, wird im o. g. Scinexx-Beitrag festgestellt. Wer weiß, wie die Wissenschaftsgeschichte verlaufen wäre, wenn Darwin Mendel gründlich studiert hätte?
Literatur
Crompton N (2019) Mendel‘sche Artbildung und die Entstehung der Arten. Internetartikel, https://www.wort-und-wissen.org/wp-content/uploads/b-19-3_mendel.pdf
Fairbanks DJ (2020) Mendel and Darwin: untangling a persistent enigma. Heredity 124, 263–273.
Podbregar N (2022) Mendel und Darwin. Wie stand der Mönch zur Evolution? Internetartikel, https://www.scinexx.de/dossierartikel/mendel-und-darwin/
Autor dieser News: Reinhard Junker
© 2022, http://www.genesisnet.info/schoepfung_evolution/n304.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
14.07.22 Gibt es doch keine neutralen Mutationen?
Mutationsforschung bringt ein wichtiges evolutionstheoretisches Konzept ins Wanken
Lange Zeit ging man davon aus, dass es aufgrund des redundanten Charakters des genetischen Codes (d. h. 64 Codons codieren für nur 20 Aminosäuren) neutrale und nicht-neutrale Mutationen in proteincodierenden Genen gibt. Auf der Grundlage dieser Annahme wurden seit über drei Jahrzehnten zahlreiche Arbeiten veröffentlicht, in denen das Verhältnis von neutralen und nicht-neutralen Mutationen als Indikator für Selektion diskutiert wurde. Eine neue Studie in der Zeitschrift Nature zeigt nun, dass es solche neutralen Mutationen wohl kaum gibt.
„Mutationen torpedieren Lehrmeinung“ titelte eine populärwissenschaftliche Webseite Juni 2022 (Podgrebar 2022). Meistens sind solche aussagekräftigen Titel übertrieben und sollen nur die Aufmerksamkeit der Leser gewinnen. In diesem Fall war der Titel aber sehr gut gewählt, da er sich auf ein seit langem bestehendes theoretisches Konzept der Evolutionsbiologie bezieht: neutrale Mutationen. Die neuen Beobachtungen, die in der Fachzeitschrift Nature publiziert wurden (Shen et al. 2022), haben weitreichende Konsequenzen für die Evolutionsbiologie – insbesondere bei der Frage, welche Rolle die Selektion in der Evolution von proteincodierenden DNA-Sequenzen spielt. Um zu verstehen, was die neuen Daten bedeuten, müssen wir uns zunächst der Genetik widmen, die den Mutationen zugrunde liegt, und wir müssen verstehen, wie Evolutionsbiologen aus dem Mutationsgeschehen die Art der Selektion ableiten. Doch zuvor schauen wir uns ganz grundlegend einmal an, wie der genetische Code funktioniert, der für die Codierung der Proteine verantwortlich ist.
Proteine sind Werkzeuge und wichtige Bausteine der Zelle und bestehen aus langen Ketten von Aminosäuren. Es gibt 20 verschiedene Aminosäuren, die eine fast unendliche Vielfalt von verschiedenen Proteinen bilden können. Man kann sich ein Protein wie eine Perlenkette vorstellen, bei der jede Perle eine bestimmte Aminosäure darstellt. In menschlichen Zellen gibt es etwa 21.000 verschiedene Proteinfamilien. Sie alle haben unterschiedliche Funktionen. Jedes einzelne Protein ist im Genom (Erbgut) codiert, durch sogenannte proteincodierende Gene, lange Abschnitte von Nukleotiden (A, C, G und T). Wie ein Protein kann man sich auch ein Gen auch wie eine Perlenkette vorstellen, bei der jede Perle ein DNA-Buchstabe ist. Abb. 1 zeigt, wie die 64 Codons des genetischen Codes mit den 20 Aminosäuren verbunden sind.
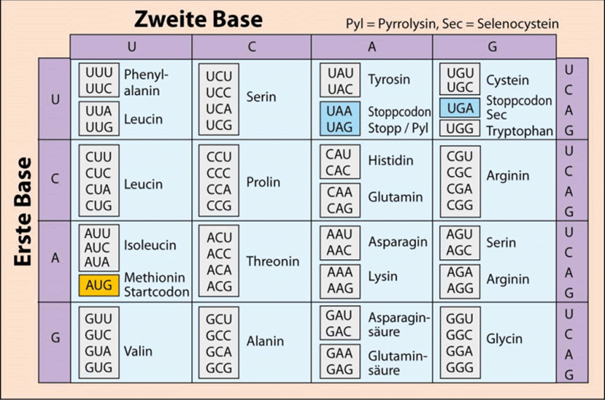
Abb. 1: Der genetische Code in seiner Standardform. Er ist hier auf der mRNA-Ebene angegeben, dort wird statt Thymin die Nukleobase Uracil (U) verwendet. Pyrrolysin (Pyl) und Selenocystein (Sec) sind selten vorkommende Aminosäuren, die als Bausteine von Proteinen erst in den letzten Jahren entdeckt wurden. In der rechten Spalte ist die dritte Base im Triplett angegeben.
Um als Information für die Gestaltung der Proteine zu fungieren, enthalten die Gene einen verschlüsselten Bauplan für Proteine: eine bestimmte Abfolge ihrer vier „Buchstaben“, der Nukleotide A, C, G und T. Diese Abfolge bestimmt die Abfolge von Aminosäuren bei der Bildung von Proteinen. Allerdings ist zu erwähnen, dass die DNA-Buchstaben des Erbgutes nicht direkt zu Proteinen umgesetzt werden, sondern dass eine Abschrift, die Messenger-RNA (mRNA) die Information vom Erbgut im Zellkern zu den Proteinfabriken, den Ribosomen, bringt (sie besitzt U statt T und ist die Abschrift eines Genabschnittes). Jeweils drei Nukleotide auf der mRNA stehen für eine bestimmte Aminosäure (und einige stehen für Start bzw. Stopp). Mit den vier RNA-Buchstaben (Nukleotide A, T, C und U) können durch Dreiergruppen insgesamt 64 verschiedene Kombinationen zusammengestellt werden. Diese sogenannten Tripletts – oder Codons – codieren also die Bausteine der Proteine (die Aminosäuren), wovon es aber nur 20 unterschiedliche in Lebewesen gibt. Es gibt also viel mehr Codons (nämlich 64) als zu codierende Proteinbausteine (20 Aminosäuren). Zusammen mit einem Codon, das als Stoppsignal funktioniert und anzeigt, wo das Protein enden muss, würden die Lebewesen theoretisch nicht mehr als 21 Codons benötigen, um alle beliebigen Proteine codieren zu können. Wozu dienen also die restlichen 43 Kombinationen des Codes?
Da es drei unterschiedliche Stoppsignale gibt, verbleiben 61 Codons, die den Satz von 20 Aminosäuren codieren, so dass viele Aminosäuren durch mehrere unterschiedliche Triplett-Kombinationen codiert sind. Den meisten Aminosäuren entsprechen also mehrere Codons. Die Aminosäuren Serin, Arginin und Leucin werden beispielsweise von jeweils sechs unterschiedlichen Codons codiert. Vier verschiedene Codons stehen jeweils für fünf andere Aminosäuren, nämlich Alanin, Glycin, Prolin, Threonin und Valin. Die übrigen Aminosäuren werden jeweils durch ein Codon, zwei oder drei Codons codiert. Aminosäuren die durch fünf Codons codiert werden, kommen nicht vor. Diese seltsame, ungleiche Verknüpfung von Codons und Aminosäuren war lange Zeit rätselhaft, hat aber damit zu tun, dass sie in verschiedener Hinsicht optimal ist, insbesondere in Bezug auf Fehlertoleranz (Freeland et al. 2000).
In der Fachsprache heißt es, der Proteincode sei ein degenerierter Code. Damit ist nicht gemeint, dass er irgendwie defekt sei, sondern dass eine bestimmte semantische Einheit (hier eine bestimmte Aminosäure) durch mehrere unterschiedliche syntaktische Abfolgen (hier verschiedene Codons) codiert wird. Da es mehrere verschiedene Codons für dieselbe Aminosäure gibt, kann man auch von redundanter Codierung sprechen.
Aufgrund dieser Redundanz gibt es sogenannte synonyme und nicht-synonyme Mutationen. Nicht-synonyme Mutationen verändern mit der Triplett-Codierung auch die Aminosäuresequenz. Da dies die Faltung (3-dimensionale Struktur der Aminosäurekette) und Funktion eines Proteins verändern kann, werden sie als nicht-neutral eingeschätzt. Synonyme Mutationen dagegen verändern zwar ebenfalls die Triplett-Codierung, jedoch nicht die Aminosäuren-Abfolge und damit auch nicht die Funktion des Proteins, da trotz Änderung die gleiche Aminosäure codiert und damit eingebaut wird. Da synonyme Mutationen keinen Einfluss auf die Sequenz und Funktion der Proteine haben, wurden sie stets als neutral betrachtet.
Verhältnis synonymer zu nicht-synonymer Mutationen: Anzeiger für Selektion?
In der Evolutionsbiologie ist das Verhältnis zwischen der Anzahl nicht-synonymer und synonymer Mutationen in homologen (gleichartigen) proteincodierenden Genen verschiedener Organismen ein häufig gebrauchter Parameter. Dieses sogenannte Ka/Ks-Verhältnis (a = nicht-synonym, s = synonym) wird verwendet, um auf die Richtung und das Ausmaß der natürlichen Selektion zu schließen, die auf proteincodierende Gene wirkt. Ein Ka/Ks-Verhältnis von mehr als 1 bedeutet positive Selektion (die Veränderungen vorantreibt); weniger als 1 bedeutet reinigende oder stabilisierende Selektion (die Veränderungen entgegenwirkt); und ein Verhältnis von genau 1 bedeutet neutrale (d. h. keine) Selektion. Viele Tausend Veröffentlichungen auf dem Gebiet der Evolutionsbiologie verwenden diese Methode, die nur unter der Annahme gültig ist, dass synonyme Mutationen (Ks) in proteincodierenden Genen tatsächlich neutral oder nahezu neutral sind.
In der Nature-Veröffentlichung von Juni 2022 wurde nun ein experimenteller Nachweis veröffentlicht, wonach die mit dem Ka/Ks-Verhältnis verbundenen Annahmen möglicherweise nicht korrekt sind, da in repräsentativen Hefegenen auch synonyme Mutationen meist eindeutig nicht-neutral sind (Shen et al. 2022).
Die Experimentatoren der Studie erzeugten mehrere Tausend Hefemutanten, die synonyme und nicht-synonyme Mutationen für 21 Gene mit unterschiedlichen Funktionen und Expressionsniveaus (d. h. wie häufig die Gene abgeschrieben werden) trugen. Anschließend maßen sie die Fitness dieser Mutationen im Vergleich zum Wildtyp. Es zeigte sich, dass 3/4 der synonymen Mutationen unerwartet zu einer signifikanten Verringerung der Fitness (d .h. der Überlebens- und Fortpflanzungsfähigkeit) führten, und „die Verteilung der Fitnesseffekte war insgesamt ähnlich – wenn auch nicht identisch – zwischen synonymen und nicht-synonymen Mutationen“ (Shen et al. 2022). Darüber hinaus fand man heraus, dass synonyme Mutationen häufig die Aktivität des mutierten Gens beeinträchtigen, indem zu viel oder zu wenig mRNA von dem Gen abgeschrieben wurde. Aus dem Ausmaß dieser Störung konnte teilweise der Fitness-Effekt vorausgesagt werden. Außerdem erwies sich die dabei gebildete Boten-RNA (mRNA) als weniger stabil. Die Autoren kommen zu dem Schluss, dass die „starke Nichtneutralität der meisten synonymen Mutationen, falls sie auch für andere Gene und Organismen gilt, eine Überprüfung zahlreicher biologischer Schlussfolgerungen über Mutation, Selektion, effektive Populationsgröße, Divergenz-Zeit und Krankheitsmechanismen, die auf der Annahme beruhen, dass synonyme Mutationen neutral sind, erforderlich machen würde“ (Shen et al. 2022).
Diese experimentelle Arbeit stellt somit ein wichtiges theoretisches Konzept in Frage, nämlich dass synonyme Mutationen in proteincodierenden Genen in jeder Hinsicht neutral sind. Darauf aufbauende evolutionstheoretische Schlussfolgerungen über Selektionswirkung und die anderen im vorigen Zitat genannten Aspekte sind damit ebenso fragwürdig. Das könnte weitreichende Konsequenzen haben. Man wird allerdings abwarten müssen, ob sich die Ergebnisse an weiteren Organismen und Genen bestätigen werden.
Zwei Dinge kann man aus den neuen Befunden jetzt schon lernen: 1. Die Vernetzungen und Zusammenhänge der Vorgänge in den Lebewesen sind viel komplexer als bisher bekannt: Mutationen haben nicht nur Einfluss auf die Abfolge der Aminosäuren der Proteine, sondern können auch andere Aspekte verändern. 2. Einmal mehr zeigt sich: Wissenschaft ist vorläufig, und das kann auch für etablierte Konzepte gelten.
Quellen
Freeland SJ, Knight RD, Landweber LF & Hurst LD LD (2000) Early Fixation of an Optimal Genetic Code. Molecular Biology and Evolution 17, 511–518. https://academic.oup.com/mbe/article/17/4/511/1127636
Podgrebar N (2022) Mutationen torpedieren Lehrmeinung. https://www.scinexx.de/news/medizin/stille-mutationen-sind-doch-schaedlich/
Shen X, Song S, Li X et al. (2022) Synonymous mutations in representative yeast genes are mostly strongly non-neutral. Nature, https://doi.org/10.1038/s41586-022-04823-w
Autor dieser News: Peter Borger
© 2022, http://www.genesisnet.info/schoepfung_evolution/n302.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
09.03.21 Die Frage nach der Entstehung von Arten – ein Zwischenruf
Die Entstehung der Arten ist mit Charles Darwins berühmtem Buch ein grundlegender Denkansatz in evolutionären Konzepten. Bei der heutigen Kenntnis molekularbiologischer Zusammenhänge könnte man meinen, dass dieser Aspekt weitgehend geklärt sei. Drei Fachleute haben in einem Beitrag auf erfrischende Weise auf die Dinge hingewiesen, die wir derzeit nicht wissen.
Drei Biologen, Evolutionsbiologen und Populationsgenetiker, haben einen Perspektiv-Beitrag in „Science“ mit der Titelfrage überschrieben: Wie viele genetische Veränderungen sind für die Entstehung neuer Arten erforderlich?“1 (Nosil, Feder & Gompert 2021). Sie eröffnen den Text mit dem Lehrbuchwissen, dass neue Arten die Vielfalt der Lebewesen bewirken und deren Neubildung oft evolutiv durch natürliche Selektion verursacht wird. Die folgende Feststellung aber lässt aufhorchen: Wie viele genetische Veränderungen zur Artbildung erforderlich sind, ist weitgehend unbekannt! Die Autoren führen verschiedene, von den Fachleuten diskutierte theoretische Modelle an. Wenn beispielsweise keine geographische Isolation im Spiel ist, erfolgt Artneubildung durch eine kleine Anzahl von Genen, was dem ausgleichenden Effekt durch den Genfluss (Vermischung) entgegenwirkt. Untersuchungen an Insekten und Pflanzen haben aber gezeigt, dass genetische Veränderungen in nur wenigen Bereichen des Genoms oft mit bestimmten Erscheinungsformen (Phänotyp) im Zusammenhang stehen und z. B. bei Schmetterlingen nur zu verschiedenen Morphen (Flügelmuster-Variationen), nicht aber zu neuen Arten führen. Nosil et al. führen auch die Buntbarsche (Cichlidae) an, in denen wenige Genbereiche die unterschiedlichen Farbmuster bestimmen. Aber neue Arten entstehen dadurch nicht. Damit meinen sie hier, dass zu einer stabilen, das Genom umfassenden Differenzierung auch eine veränderte Kiefergestalt und Körperform einbezogen werden müssen; dass also viele genetische Bereiche betroffen sind. Am Ende der Diskussion kommen die Autoren zu dem Schluss, dass bei der Artneubildung wenige oder viele Genbereiche involviert sein können; es komme dabei auf die Umstände an.
Die Autoren fügen an, dass noch viel Arbeit zu tun bleibt, um experimentell nach den ursächlichen Effekten der verschiedenen Einflussgrößen der Artbildung zu suchen. Sie äußern die Hoffnung, dass Evolutionsbiologen durch die Vereinigung von Theorie und empirischen Daten eine Möglichkeit haben, die Entstehung neuer Arten besser zu verstehen. Nosil et al. beschließen ihren Beitrag mit der Feststellung, dass eine grundsätzliche Hauptfrage das Ausmaß betreffe, in dem mikroevolutive Prozesse größere, makroevolutive Muster der Biodiversität erklären können, wie dies bei der Ausbreitung der Buntbarsche (Cichlidae) beobachtet werden kann.
Zunächst ist es erfrischend zu lesen, wie viel Unwissen hinter etablierten Lehrbuchaussagen stecken kann; das kann zu Freiraum für ungebundeneres Nachdenken führen. Für den Autor war der Schlusssatz insofern sehr aufschlussreich, als für die Vielfalt der Familie der Buntbarsche (Cichlidae) auf der chromosomalen Ebene durch die Mendel‘schen Gesetze und auf der molekularbiologischen Ebene durch die transponierbaren Elemente, durchaus erfolgversprechende Erklärungsansätze vorliegen (Crompton 2019). Evolutionsbiologen aber haben noch eine viel größere Aufgabe zu lösen, nämlich wie die Fülle der unterschiedlichsten Baupläne der Lebewesen entstanden sein kann. Verglichen damit sind die Herausforderungen für diejenigen, die von einer Schöpfung ausgehen, bei diesem Thema relativ klein!
Literatur
Crompton N (2019) Mendel´sche Artbildung und die Entstehung der Arten. 1. Woher kommt die Artenvielfalt? Stud. Integr. J. 26, 86-92.
Nosil P, Feder JL & Gompert Z (2021) How many genetic chances create new species? Science 371, 777-779.
Anmerkung
1 “How many genetic chances create new species?“
Autor dieser News: Harald Binder
© 2021, http://www.genesisnet.info/schoepfung_evolution/n288.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
08.01.21 „Überraschungen der Evolution“
Evolutionäre Neuheiten entstanden als Nebeneffekte evolutionärer Veränderungen durch Neuverwendung schon vorhandener Bauteile. Auf genetischer Ebene teilen Arten ganz verschiedener Tierstämme einen ähnlichen „Baukasten“. Dies ermögliche als Frucht der Evolutionsforschung auch medizinischen Fortschritt. So sieht es der Paläontologe Neil Shubin in einem Webinar-Beitrag. Tatsächlich sind die Befunde evolutionstheoretisch gar nicht leicht zu verstehen, lassen sich aber im Rahmen von Schöpfung befriedigend deuten.
Unter der Überschrift „Evolutions’s surprises“ fasste Sara Frueh von der National Academy of Sciences (USA) einen Vortrag des Paläontologen Neil Shubin zusammen, den dieser bei einem Webinar im April 2020 gehalten hat (Frueh 2020). Die „Überraschungen“ beziehen sich auf neuere evolutionstheoretische Vorstellungen.
Shubin findet es überraschend, dass die Veränderungen, die zu Schlüsselereignissen der Evolution wurden – wie die Eroberung des Landes ausgehend von Fischen, oder die Eroberung der Luft durch die Vögel – lange vor diesen Meilensteinen erfolgten und auch zu anderen Zwecken. So seien Lungen nicht entstanden, als die Tiere im Begriff waren, an Land zu gehen. Ebenso seien die Federn nicht entstanden, als die Tiere im Begriff waren, zu fliegen. Lungen bzw. Federn waren vielmehr in Formen ausgebildet, die „noch“ im Wasser bzw. noch an Land lebten – und eben nicht, um den Landgang bzw. den Flug zu ermöglichen. Vielmehr habe es Lungen und die Vierbeiner-Extremität bereits bei wasserlebenden Formen gegeben. Diese Merkmale seien nur „umfunktioniert“ worden, als es nötig war. Die Lungen hätten Fischen ermöglicht, Luft von der Wasseroberfläche zu schlucken, wenn der Sauerstoffgehalt zu niedrig war, um genügend davon durch die Kiemen zu erhalten. Die schon vorhandenen Lungen konnten dann beim Landgang verwendet werden. Und der von Shubin und seinem Team entdeckte Tiktaalik roseae besaß landtierartige Gliedmaßen, obwohl er wie ein Fisch im Wasser lebte. Ähnlich wird bei vogeltypischen Merkmalen argumentiert: Beispielsweise hätten sich Federn vermutlich entwickelt, um die Körpertemperatur der Dinosaurier zu regulieren, oder vielleicht zur Färbung und Balz. Später seien sie als Teil des Flugapparats verwendbar gewesen. „Es ging darum, alte Erfindungen auf neue Art und Weise für neue Zwecke zu nutzen“, so Shubin. „Das ist es, worum es in der Evolution geht.“

Abb. 1: Nachbildung des Skelettfundes von Tiktaalik roseae. (Wikimedia: Tiktaalik_belgium.JPG: Esv derivative work: Petter Bøckman (talk) – Tiktaalik_belgium.JPG, CC BY-SA 3.0)
Indirekter Evolutionsweg ohne Ziel? Die Einschätzung von Shubin ist doppelt interessant. Zum einen: Die hypothetische evolutive Entstehung von Schlüsselmerkmalen soll nicht direkt erfolgen, also nicht so, als ob diese direkt angesteuert würden. Klar, möchte man sagen, Evolution erfolgt nicht zielorientiert. Evolutionstheoretiker werden nicht müde, das zu betonen. Zielorientierung ist nur möglich, wenn man einen Schöpfer annimmt, doch diese Möglichkeit wird in der Regel ausgeschlossen. Also müssen scheinbare Ziele so erreicht werden, als hätte man nie darauf abgezielt, selbst wenn es sich wie bei den Vierbeinern und den Vögeln um die Eroberung eines ganz neuen Lebensraums geht.
Zum anderen kommt Shubin aber auch bei der Beschreibung eines evolutiven Umweges bzw. evolutiver Vorstufen nicht ohne die Zweckkategorie aus. „Wozu“ sind die Einrichtungen für den Landgang bzw. für das Fliegen entstanden, wenn nicht für die Eroberung des Landes bzw. der Luft? „Zu anderen Zwecken“, sagt Shubin, was doch wieder eine Zielvorgabe impliziert. Evolutionstheoretiker haben sich seit Darwin jedoch zum Ziel gesetzt, Evolutionsmechanismen als rein natürliche Prozesse zu beschreiben. Die offensichtliche Zweckmäßigkeit der Strukturen des Lebens holt die Biologen jedoch immer wieder ein. Um eine bestimmte Zielorientierung (z. B. fürs Fliegen; „for fly“) zu vermeiden, weicht man auf ein anderes Ziel aus (z. B. Regulation der Körpertemperatur oder die Balz).
Sind die Szenarien plausibel? Der Grundgedanke von Shubin ist nicht neu. Schon im Biologiestudium konnte man vor über 50 Jahre lernen, dass Neuheiten in der Evolution auftreten würden, weil es „Voranpassungen“ (Präadaptionen) gebe – wobei „Anpassung“ wiederum ein teleologischer Begriff ist (also ein Begriff, der ein Ziel beinhaltet). Aber davon abgesehen – hilft dieses Konzept weiter? Vielleicht ein bisschen, aber nicht, um die entscheidenden Veränderungen zu erklären. Wenn beispielsweise Federn ursprünglich der Thermoregulation oder der Balz dienten, haben solche „Federn“ damit noch praktisch keine Eigenschaften, die zum Fliegen verhelfen. Die eigentliche Herausforderung, nämlich die zahlreichen fürs Fliegen erforderlichen anspruchsvollen Eigenschaften zu erwerben, wäre evolutiv erst noch zu meistern (genauer erläutert wird das in Junker [2017]). Ähnlich ist die Problematik bei der Entstehung der Fortbewegung auf dem Land. Es sind ganz andere Anforderungen zu bewältigen, um sich mit einer Vierbeiner-Extremität auf dem Land fortzubewegen, als wenn man das im Wasser tut bzw. – evolutionär betrachtet – wenn man das bisher nur im Wasser gemacht hat.
Wie kann man die Befunde interpretieren, wenn man von Schöpfung ausgeht? Shubin stellt fest, dass Merkmale, die typisch für eine bestimmte Lebensweise sind (Leben auf dem Land oder in der Luft), in teils ähnlicher Form auch bei Arten vorkommen, die andere Lebensräume besiedeln. Es gibt landlebende Dinosaurier mit haarartiger Körperbedeckung (die man nicht unbedingt als „Federn“ bezeichnen muss), mit hohlen Knochen und diversen weiteren vogeltypischen Merkmalen. Und es gibt Fische mit Fingern und mit einfachen Lungen. Die Merkmale sind jedoch auf die verschiedenen Arten bzw. Gattungen häufig so unsystematisch verteilt, dass sie keine geradlinige Evolution belegen, sondern in unterschiedlichen Kombinationen auftreten. Dies führt dazu, dass keine widerspruchsfreien Baumschemata konstruiert werden können, sondern viele Konvergenzen angenommen werden müssen. Sehr ausgeprägt ist das bei den Theropoden-Dinosauriern der Fall, die in die Nähe der Vögel gestellt werden (Junker 2019).
Shubin spricht von einem „Baukasten“ auf genetischer Ebene; und es scheint, dass auch auf morphologischer Ebene eine Art Baukastensystem erkennbar ist. Was die Gene betrifft, sagt Shubin: „Der grundlegende Werkzeugkasten, der die Körper von Lebewesen wie uns und Mäusen aufbaut, wird mit Würmern, Fliegen und Fischen geteilt.“ Und er deutet diesen Befund evolutionstheoretisch so: „Alle Werkzeuge, die zum Aufbau unseres Körpers verwendet werden, sind ursprünglich in Fliegen und anderen Lebewesen entstanden.“ Ein Baukastensystem ist aber im Rahmen eines Schöpfungsparadigmas leicht zu verstehen. Denn ein Schöpfer ist in seinem Handeln frei; er kann Merkmale erschaffen und kombinieren, wie er will; nur sollten die Lebewesen überlebensfähig sein. Dagegen ist Evolution an einen natürlichen, nicht-zielgerichteten Mechanismus gebunden. Wie aber kommt es ohne Zielvorgabe durch rein natürliche Mechanismen unabhängig immer wieder zu ähnlichen Bauelementen? Und wie wurde der Baukasten zu Beginn zusammengestellt?
Anwendung in der Medizin? Da es eine Art Baukasten auf genetischer Ebene gibt und die Arten verschiedenster Tierstämme daran teilhaben, sieht Shubin positive Konsequenzen aus der evolutionsbiologischen Forschung für die Medizin. „Diese Verbindungen ermöglichen wissenschaftliche und medizinische Entdeckungen, die letztlich auch dem Menschen helfen können“, erklärt Shubin. Denn wenn man versteht, wie Gene in Tieren bei Gesundheit und Krankheit an- und abgeschaltet werden, lerne man medizinisch relevante Dinge auch über den Menschen, bei dem dieselben Gene vorkommen. Doch das hat an sich mit Evolution nichts zu tun, sondern – sofern die Übertragung der Erkenntnisse auf den Menschen wirklich tragfähig ist – mit dem Baukastensystem, das, wie erwähnt, aus der Sicht der Schöpfungslehre plausibler erscheint und leichter verstehbar ist als aus der Sicht einer ziellos verlaufenden Evolution.
Anmerkung
Originalzitate unter https://www.nationalacademies.org/news/2020/08/evolutions-surprises
Quellen
Frueh S (2020) Evolution’s Surprises. https://www.nationalacademies.org/news/2020/08/evolutions-surprises
Junker R (2017) Dino-Federvieh – Zum Ursprung von Vogelfeder und Vogelflug. https://wort-und-wissen.org/wp-content/uploads/b-17-1_feder-und-flug.pdf
Junker R (2019) Sind Vögel Dinosaurier? Eine kritische Analyse fossiler Befunde. https://wort-und-wissen.org/wp-content/uploads/b-19-4_dinos-voegel.pdf
Autor dieser News: Reinhard Junker
© 2021, http://www.genesisnet.info/schoepfung_evolution/n284.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
01.10.19 Gregor Mendel, die Entstehung der Arten und Schöpfung
Der Augustinermönch Gregor Mendel ist für seine Vererbungsgesetze bekannt. Kaum jemand weiß jedoch, dass Mendel ein Konzept entwickelt hat, das eine Erklärung der Entstehung einer großen Formenvielfalt innerhalb genetischer Familien (Grundtypen) innerhalb weniger Generationen ermöglicht. Dieses Konzept erweist sich gerade angesichts aktueller Befunde als ausgesprochem fruchtbar. Zugleich ist es für die Modellierungen im Rahmen der Schöpfungslehre von besonderem Interesse.
Die Frage, wie Artbildung erfolgt, also die Aufspaltung von einer Art in zwei oder mehr Tochterarten, gehört zu den zentralen Fragen der Biologie. Charles Darwins epochemachendes Werk „On the Origin of Species“ trägt diese Frage implizit im Titel. Im Gefolge Darwins sollen Arten sich dadurch auseinanderentwickeln und aufspalten, dass sich nach und nach Mutationen ansammeln, Populationen geographisch getrennt werden und in den getrennten Populationen unterschiedliche Selektion wirkt. Im Laufe der Zeit sollen schließlich Fortpflanzungsbarrieren entstehen (genetische Isolation). Auf dieser Basis werden große Zeiträume für die Bildung neuer Arten veranschlagt. Dieses Verständnis ist heute Mainstream in der Evolutionsbiologie.
Ein Zeitgenosse Darwins, der Augustinermönch Gregor Mendel, der als Vater der modernen Genetik bezeichnet wird, verfolgte ein ganz anderes Konzept. Dieses ist weitgehend übersehen worden, erweist sich aber gerade angesichts der Befunde über sehr schnelle Artbildungsprozesse als ausgesprochen aktuell und fruchtbar. Es beruht auf präexistenter Vielfalt durch präexistente genetische Programme und ermöglicht Artbildung innerhalb weniger Generationen, da die Unterschiede zwischen den Arten nicht in langwierigen Mutations-Selektions-Zyklen aufgebaut werden müssen, sondern von Beginn an bereits angelegt sind. Mendels Gesetz der Kombinationen von Merkmalsausprägungen erweist sich angesichts zahlreicher neuerer Befunde als ausgesprochen nützlich zum Verständnis der Veränderung von Arten innerhalb genetischer Familien.
Nigel Crompton, Biologe an der Cornerstone-University (Michigan/USA; https://www.cornerstone.edu/staff/nigel-crompton) hat sich mit Mendels Konzept und Mendel’scher Artbildung intensiv befasst und seine Ergebnisse in einem Internet-Artikel der Studiengemeinschaft Wort und Wissen publiziert. Dieses Konzept erweist sich angesichts einer zunehmenden Anzahl von Beispielen schneller Artaufspaltungen innerhalb von Grundtypen als sehr fruchtbar: Die Formenvielfalt innerhalb von Grundtypen „lebt“ von latenten (= verborgenen), präexistenten Variationsprogrammen, die im Laufe von Generationen durch Homozygotisierung (= Reinerbig-Werden), durch Wechselwirkung zwischen verschiedenen Genen (Epistasis) und durch Springen von Genen ausgeprägt werden. Die heute vorliegenden Befunde sind damit besser in einem Schöpfungsrahmen verstehbar als im Rahmen neodarwinistischer Modellierungen.
Zusammenfassung von Cromptons Arbeit
„Mendels bahnbrechende wissenschaftliche Abhandlung aus dem Jahre 1866, Versuche über Pflanzenhybriden, ist voller Überraschungen und tiefer Einsichten. In ihr beschreibt er Eigenschaften von Hybriden, die sowohl dominante als auch rezessive Merkmalsausprägungen haben, und legt dar, dass rezessive Merkmalsausprägungen in den Hybriden latent vorhanden sind. Er beschreibt, wie diese latenten Merkmalsausprägungen exprimiert werden und neue Sorten und Arten von Hülsenfrüchten erzeugen können. Mendels Entdeckungen lassen die genetische Strategie hinter der Entstehung der Arten (innerhalb von separaten Familien) erkennen. Sein Gesetz der Kombinationen von Merkmalsausprägungen resultiert durch meiotische Rekombination.
Eigenartigerweise wird dieses Mendel‘sche Gesetz von Biologen meistens übersehen oder bewusst ignoriert. Dabei lässt sich mit Hilfe dieses Gesetzes die Entstehung einer großen Anzahl von phänotypischen Variationen zufriedenstellend erklären. Wenn alternative Kombinationen von Merkmalsausprägungen in den Nachkommen fixiert werden, kann daraus eine Fülle von neuen Arten und Gattungen entstehen. Die für diese (neuen) Arten notwendige Information ist bereits vorhanden, aber sie befindet sich latent in den Geschlechtszellen, und zwar in ihrer DNA, wie wir heute wissen. Die Information wird durch Prozesse wie Dominanz, Epistasis* und Transposition* in einem latenten Zustand gehalten, wie man das bei einigen der von Mendel untersuchten Merkmale feststellen kann. Aus der Rekombination dieser Information ergeben sich Familien von Organismen. Die im pan-heterozygoten* Genom* einer Vorfahrenpopulation vorhandene Information erfährt aufeinander folgende Meiosen* und lässt dadurch verschiedene Kombinationen von homozygoten* dominanten und rezessiven Merkmalen entstehen, aus denen dann die zahlreichen Arten und Gattungen entstehen, die zusammen eine separate Familie bilden. Wenn neue Kombinationen von homozygoten Merkmalen auftreten, sorgt die Fortpflanzungsisolation dafür, dass sie fixiert bleiben. Die Selektion ihrerseits begünstigt in jeder einzelnen Situation lediglich die geeignetste dieser Kombinationen.
*Worterklärungen:
- Epistasis (= Wechselwirkung zwischen Genen)
- Transposition (= Springen von Genen an neue Positionen im Erbgut)
- pan-heterozygot (= umfassend mischerbig)
- Genom (= Erbgut)
- Meiose (= Reifeteilung von Geschlechtszellen)
- homozygot (= reinerbig)
Es stellt sich heraus, dass Mutationen zwar einen zusätzlichen Beitrag zur phänotypischen Vielfalt der Arten leisten, jedoch nicht notwendig sind, um die umfangreichen Merkmalsunterschiede hervorzubringen, die man innerhalb der Arten beobachten kann. Für die Entstehung der phänotypischen Artenvielfalt von Familien spielen sie keine notwendige Rolle. Wallace und Darwin hatten zwar Variation und Anpassung beobachtet; da sie aber die Entdeckungen Mendels und die ihnen zugrunde liegenden Mechanismen nicht kannten, kamen sie zu dem Trugschluss, es könne grenzenlose Variationsmöglichkeiten geben. Neue Kombinationen von Merkmalsausprägungen spiegeln die Ausprägung zuvor schon latent vorhandener Information wider. Die Summe aller dieser Kombinationen stellt die Grenzen dar, innerhalb derer eine Familie sich durch Ausprägung der angelegten Möglichkeiten entfalten kann. Diese intrafamiliäre oder cis-Evolution erzeugt einen separaten Abstammungsbaum des Lebens. Mehrere solcher separater Familien (Bäume) ergeben einen Wald des Lebens. Die Entdeckungen Mendels, die er in seiner faszinierenden wissenschaftlichen Abhandlung wiedergibt, waren die Geburtsstunde der Wissenschaftsdisziplin der Genetik und zeigten, woher neue Arten rasch und auch lebenstüchtig entstehen können.“
Der komplette Artikel (32 Seiten, PDF) kann hier heruntergeladen werden:
http://www.wort-und-wissen.de/artikel/sp/b-19-3_mendel.pdf
Autor dieser News: Reinhard Junker
© 2019, http://www.genesisnet.info/schoepfung_evolution/n270.php
09.12.14 „Brauchen wir eine neue Evolutionstheorie?“
Unter der Fragestellung „Does evolutionary theory need a rethink?“ erschien Anfang Oktober in der Wissenschaftszeitschrift Nature ein Pro und Contra zweier Gruppen von Wissenschaftlern. Ende November wurde eine deutsche Übersetzung veröffentlicht.1 Die Auseinandersetzung zeigt, dass es nicht die eine, alles erklärende Evolutionstheorie gibt, sondern eine Vielzahl von theoretischen Ansätzen, die zum Teil miteinander konkurrieren oder gar einander ausschließen. Manche Ansätze laufen darauf hinaus, dass eine Zielorientierung angenommen werden muss. Das erinnert an den Design-Ansatz, doch die Quellen für Zielsetzungen werden in den Lebewesen gesucht, während ein willensbegabter geistiger Urheber tabu ist.
Als im Jahr 1959 das hundertjährige Jubiläum von Charles Darwins Evolutionstheorie gefeiert wurde, galten die wesentlichen Fragen der Evolutionsforschung als geklärt. Dazu gehörte auch der auf Darwin zurückgehende Mechanismus von Variation und Selektion (natürliche Auslese), der als rein natürlicher Prozess verstanden wird und gleichsam naturgesetzmäßig verlaufen soll. Die Variationen innerhalb von Arten entstehen demnach durch Mutationen, das sind Änderungen des Erbguts, die richtungslos in Bezug auf potenzielle Ziele verlaufen (und in diesem Sinne zufällig sind). Die durch Mutationen entstandenen Varianten bilden das Rohmaterial, das der Selektion zur Verfügung steht. Bei Überproduktion der Nachkommen gelangt tendenziell nur der bestangepasste Teil selbst zur Fortpflanzung und kann seine Merkmalsausprägungen an die nächste Generation weitergeben. Dieses Wechselspiel von Zufallsvariation (Mutation) und Auslese (Selektion) soll im Laufe großer Zeiträume dafür sorgen, dass Arten sich nicht nur an wechselnde Umweltbedingungen anpassen, sondern letztlich auch neue Baupläne mit völlig neuen Bauelementen entstehen.2 Seit den 1930er und 1940er Jahren hatte sich die Synthetische Evolutionstheorie etabliert und war viele Jahrzehnte ziemlich unangefochten. Deren Szenario war weitgehend Konsens im Jahr 1959 und im Großen und Ganzen bestimmt diese Sicht die öffentliche Darstellung des evolutionären Prozesses bis in unsere Zeit.
Doch diese Situation hat sich mittlerweile grundlegend geändert. Eine Fülle neuer Ansätze für kausale Evolutionstheorien wurde in die Diskussion eingebracht und viele Fachartikel beginnen mit Feststellungen wie: „… we have made relatively little progress in understanding how novel traits come into being in the first place“ (Moczek 2008, 432) oder: „How body pattern evolves in nature remains largely unknown“ (Cleves et al. 2014, 13912). Diese Einschätzungen überraschen, da doch schon vor über 50 Jahren häufig zu lesen und zu hören war, die Frage nach den Evolutionsmechanismen sei im Wesentlichen geklärt. Offenbar ist einer wachsenden Zahl von Biologen bewusst geworden, dass mit den einst als vollständig erachteten kausalen Faktoren der Synthetischen Theorie nichts wirklich Neues zustande kommt. Evolution, als naturgeschichtlicher Prozess, aber gilt dessen ungeachtet als nicht hinterfragbare Tatsache. Also muss nach neuen Wegen gesucht werden, auf denen u. a. die Entstehung neuer Baupläne verlaufen konnte.
Eine Gruppe von Evolutionsbiologen sieht nach wie vor keinen Grund zu einem grundsätzlichen Umdenken in der Frage nach den Triebfedern und Mechanismen der Evolution. Andere dagegen fordern eine deutliche Erweiterung bisheriger Theorien. Mittlerweile haben sich in dieser Frage verschiedene Lager gebildet. Dieser Eindruck wird bestätigt, wenn man ein Pro und Contra verfolgt, dass die Wissenschaftszeitschrift Nature Anfang Oktober veröffentlicht hat (Laland et al. 2014, Wray et al. 2014). Eine Gruppe, vertreten durch acht Wissenschaftler meint, die Evolutionstheorie brauche dringend ein Umdenken, es sei eine Erweitere Evolutionäre Synthese (EES) unter Einbeziehung bisher vernachlässigter Disziplinen erforderlich (Pro-Gruppe). Das sei kein Sturm im Wasserglas, sondern ein Kampf um die Seele der Evolution. Sieben Wissenschaftler, die die traditionelle Sicht vertreten (Contra-Gruppe), halten dagegen: Mit der (kausalen) Evolutionstheorie sei alles in Ordnung. Das verfügbare Wissen, dass die Pro-Gruppe einfordere, sei schon berücksichtigt oder ließe sich auch in Zukunft in den bekannten Rahmen einfügen. Es gebe keinen Anlass für eine Neukonzeption.
Vier Bausteine für eine Erweiterte Evolutionäre Synthese
Die Pro-Gruppe nennt vier Aspekte, die nach ihrer Auffassung in der Standard-Evolutionstheorie nicht als Evolutionsfaktoren berücksichtigt, sondern als deren bloße Folge angesehen würden:
- Der genzentrierte Ansatz (Genänderungen als Initialzündungen) müsse ergänzt werden durch die Berücksichtigung zahlreicher Wechselwirkungen mit äußeren und inneren Einflüssen während der ontogenetischen Entwicklung. Diese führen u. a. zu Einschränkungen der Entwicklungsrichtungen („developmental bias“, Entwicklungszwänge) und begrenzen damit mögliche Variationen von Merkmalsausprägungen ein Stück weit (es erfolgt quasi eine Kanalisierung), bevor die Umweltselektion wirkt.
- Die Umweltbedingungen haben nicht nur eine passive Rolle als Selektionsfaktoren, vielmehr werde die Umwelt durch die Lebewesen aktiv mitgestaltet (Nischenkonstruktion); dadurch beeinflussen die Lebewesen selbst auch ihre eigene Evolution.
- Durch die Plastizität der Lebewesen (Änderungen infolge von Umweltreizen – ohne Genänderungen!) sei eine schnelle Anpassung und sogar die Offenlegung bisher verborgener Merkmale möglich, die nachfolgend durch Genvariationen (Mutationen) dauerhaft sichtbar fixiert werden können.
- Extragenetische Veränderungen (Epigenetik) in der Gen-Regulation könnten wie die Gene selber ebenfalls vererbt werden und Einfluss auf Evolution nehmen.
Die Contra-Fraktion hält dagegen, dass die genannten vier Phänomene zwar tatsächlich bedeutende evolutionäre Prozesse und Gegenstand ihrer eigenen Forschungen seien, dass sie aber nicht dermaßen in den Fokus gerückt werden müssten, dass die Einführung einer neuen Bezeichnung wie „Erweiterte Synthese in der Evolutionstheorie“ gerechtfertigt sei. Es treffe nicht zu, dass diese Faktoren in der Standard-Evolutionstheorie vernachlässigt würden. Vielmehr gebe es nur neue Namen für alte Konzepte. Von der Standard-Evolution werde durch die Vertreter der EES eine Karikatur gezeichnet. Weiter sei es verfehlt, eine Gen-Zentriertheit zu kritisieren, denn Veränderungen im Erbgut hätten einen wesentlichen Anteil an Anpassung und Artbildung, was in vielen Fällen auch genau belegt sei (z. B. Antibiotikaresistenzen oder Laktosetoleranz beim Menschen). Es sei außerdem nicht geklärt, ob die Plastizität genetische Variationen im Rahmen des Adaptationsprozesses tatsächlich steuern bzw. fixieren kann. Ebenso sei die Rolle der Entwicklungszwänge (developmental bias) auf die Evolution adaptiver Merkmale schwer zu bestimmen. Schließlich gebe es keine stichhaltigen Beweise für eine tragende Rolle vererbter, epigenetischer Modifikationen auf evolutionäre Anpassung. In den vier o. g. Phänomenen sieht die Contra-Gruppe daher lediglich Erweiterungen der bislang favorisierten zentralen Prozesse des evolutionären Wandels (natürliche Selektion, Drift, Mutation, Rekombination und Genfluss). Die neu eingeforderten Mechanismen seien allesamt nicht essentiell für die Evolution, sondern modifizieren lediglich die bekannten Erklärungen für evolutionäre Prozesse unter gewissen Umständen.
Kommentar
Diese Auseinandersetzung zeigt, dass es entgegen vieler öffentlicher Verlautbarungen nicht die eine, alles erklärende Evolutionstheorie gibt, sondern eine Vielzahl von theoretischen Ansätzen, die zum Teil miteinander konkurrieren oder gar einander ausschließen (vgl. dazu Ullrich 2010). Diese Situation resultiert aus dem wissenschaftlichen Erkenntnisprozess und dokumentiert an sich einen normalen innerwissenschaftlichen Prozess. Die Brisanz ergibt sich in diesem Fall aus dem Forschungsgegenstand Evolution. Evolution wird einerseits als historisches Faktum vorausgesetzt, konnte andererseits aber auch über 150 Jahre nach Darwin bisher kausal nicht erschlossen werden. Eine Reihe von Wissenschaftlern hält beim gegenwärtigen Wissensstand die Standard-Evolutionstheorie für so ungenügend, dass sie eine substanzielle Erweiterung für erforderlich halten. Die Frage nach den Mechanismen erscheint weniger klar denn je. Die Vielfalt neuer Ansätze ist genau dafür ein gewichtiges Symptom. Es wäre wichtig, wenn dieser Aspekt stärker auch ins öffentliche Bewusstsein treten und bei der Debatte um Schöpfung und Evolution Berücksichtigung finden würde.
Ob die Lösungsansätze der Pro-Gruppe erfolgversprechend sind, ist bislang zweifelhaft, und zwar aus folgenden Gründen. Zwei der vorgeschlagenen Ansätze – die Berücksichtigung der Plastizität und der epigenetischen Vererbung für Erklärungen evolutionärer Innovationen – ermöglichen für sich alleine gar keine Evolution, da nach bisheriger Kenntnis die betreffenden Änderungen jederzeit rückgängig gemacht werden können und in der Regel nicht stabil (oder gar nicht) an die nächsten Generationen weitergegeben werden. In beiden Fällen handelt es sich um Änderungen, die zunächst in irgendeiner Form – ausgelöst durch Umweltreize – abgerufen werden und keine Änderungen im Erbgut beinhalten. Bei erneuten Änderungen dieser Umweltreize ändern sich aber auch die betreffenden Merkmalsausprägungen und fallen z. B. in den vorherigen Zustand zurück. Was ist dann für die Evolution gewonnen? Aufgrund von Umweltreizen oder epigenetisch gesteuerte Veränderungen müssen also nachfolgende Merkmalsvarianten durch passende Mutationen gleichsam festgehalten (fixiert) werden, und diese müssen dann sich in „klassischer Weise“ in den Populationen durchsetzen. Die Kritik der Contra-Gruppe ist an dieser Stelle daher verständlich. Die Mutmaßung, dass durch plastische Reaktionen konstruktive Neuheiten entstehen können, ist experimentell nicht nachvollzogen und auch theoretisch sehr unwahrscheinlich. Eine Zusammenfassung der Ansätze findet sich bei Junker (2014). Beide Faktoren – Plastizität und Epigenetik – sind aber aus einer Schöpfungsperspektive sehr interessant, ermöglichen sie doch den Lebewesen eine weitreichende Anpassungsfähigkeit und enorme Flexibilität bei wechselnden Umweltbedingungen (Näheres bei Junker 2014).
Dass ontogenetische Entwicklungszwänge evolutionär weiterhelfen sollen, ist ebenfalls fragwürdig. Denn Entwicklungszwänge und kanalisierende Randbedingungen während der Ontogenese „filtern“ zwar einerseits unpassende bzw. inkompatible Veränderungen aus, so dass nur ein Teil der potentiell möglichen Veränderungen eine Ausprägung im Erwachsenenalter erfährt – damit erfolgt eine interne Vorselektion der neuen Varianten. Aber weshalb soll andererseits dies dazu beitragen, dass neue Formen entstehen? Durch den Verweis auf ontogenetische Entwicklungszwänge wird nur ein problematischer Aspekt der Standard-Evolutionstheorie entschärft, nämlich die Zufälligkeit der Veränderungen des der Evolution zur Verfügung stehenden „Rohmaterials“. In dieser Zufälligkeit wird offenbar ein Problem gesehen, aber die vorgeschlagenen Lösungen helfen nicht in Bezug auf die Entstehung von Neuheiten. Ähnliche Kritik kann man beim Faktor „Nischenkonstruktion“ formulieren.
Design in der Natur
Bemerkenswert ist im Beitrag der Pro-Gruppe ein Satz über den Design-Ansatz („Intelligent Design“). Die Autoren mutmaßen, dass „die Evolutionsbiologen auch – gejagt vom Schreckgespenst des Intelligent Design – eine geschlossene Front gegenüber wissenschaftsfeindlichen Ansätzen bilden“ möchten. Diese Befürchtung ist berechtigt (nicht aber die Unterstellung der Wissenschaftsfeindlichkeit). Denn die Mängel der Standard-Evolutionstheorie begünstigen eben auch ganz alternative Ansätze wie den Design-Ansatz und zwar genau aus dem Grund, weshalb die Pro-Gruppe nach neuen naturalistischen Ansätzen sucht: Es braucht eine wie auch immer geartete Steuerung: Plastizität, Epigenetik, Entwicklungszwänge und Nischenkonstruktion haben nämlich in den Augen der Pro-Gruppe eines gemeinsam: Man sieht Bedarf an der Eindämmung des Zufalls und an gezielter Anpassung; die Umweltselektion wird dafür als nicht ausreichend erachtet. Im Grunde genommen werden mit diesen Ansätzen Zielorientierungen eingeführt; die sich aus dem Organismus selbst heraus ergeben sollen. Das aber stützt eine Art Design-Ansatz, mit der paradoxen Besonderheit, auf eine intentionale (willensbegabte) geistige Quelle zu verzichten. Die Integration einer intentionalen geistigen Quelle als Erklärungsoption für das Sosein des Lebens ist im wissenschaftlichen Mainstream nämlich tabu. Man wird also weiterhin natürliche Mechanismen suchen und auf den Prüfstand stellen, um rein natürliche, blinde, nichtgeistige Naturprozesse aufzuspüren, die kreativ sein sollen. Der Naturwissenschaft wird diese Vorgehensweise sicherlich weitere und unerwartete Erkenntnisse liefern. Ob diese uns jedoch der Antwort nach den Ursachen und Mechanismen der hypothetischen Evolution näher bringen, darf mit Blick auf die Geschichte bezweifelt werden.
Die Frage nach einem Schöpfer stellt sich also auch weiterhin, nicht weil die Naturwissenschaft versagt, sondern weil zu erwarten ist, dass der durch sie generierte Wissenszuwachs weiterhin Hinweise auf die Existenz einer geistigen Verursachung des Lebens liefern wird.
Quellen
Cleves PA, Ellis NA, Jimenez MT, Nunez SM, Schluter D, Kingsley DM & Miller CT (2014) Evolved tooth gain in sticklebacks is associated with a cis-regulatory allele of Bmp6. Proc. Natl. Acad. Sci. 111, 13912-13917.
Junker R (2014) Plastizität der Lebewesen: Baustein der Makroevolution? http://www.wort-und-wissen.de/artikel/sp/b-14-2-plastizitaet.pdf
Laland K et al. (2014) Does evolutionary theory need a rethink? Yes, urgently. Nature 514, 161-164.
Moczek AP (2008) On the origins of novelty in development and evolution. BioEssays 30, 432-447.
Ullrich H (2010) Evolution und Evolutionstheorien. Irrtümliche Selbstverständnisse und Fehldarstellungen naturalistischer Ursprungsmodelle. Stud. Integr. J. 17, 76-87.
Wray G et al. (2014) Does evolutionary theory need a rethink? No, all is well. Nature 514, 161-164.
Anmerkungen
1 http://www.spektrum.de/news/brauchen-wir-eine-neue-evolutionstheorie/1320620
2 „Das Endresultat der natürlichen Selektion ist es, dass jede Kreatur dahin strebt, im Verhältnis zu den Bedingungen, in denen sie lebt, immer vollkommener zu werden. Diese Verbesserung führt unvermeidlich zu einem Fortschritt in der Organisation der meisten Lebewesen“ (Charles Darwin: Über den Ursprung der Arten. London, 1859).
Autor dieser News: Reinhard Junker
© 2014, http://www.genesisnet.info/schoepfung_evolution/n221.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
19.06.17 Spannendes Buch über die grundlegende Krise von Evolutionstheorien
In seinem im Jahr 2016 erschienenen Buch „Evolution: Still a theory in crisis“ formuliert der Biochemiker Michael Denton grundlegende Kritik an (neo-)darwinistischen Evolutionstheorien und untermauert sie mit einer großen Anzahl von verschiedensten Beispielen. Seine Kritik spielt dem Schöpfungsgedanken in die Hände.
30 Jahre nach der Veröffentlichung seines vielbeachteten Werks „Evolution: A theory in crisis“ veröffentlichte der britisch-australische Biochemiker Michael Denton ein Nachfolgewerk: „Evolution: Still a theory in crisis“ (Seattle: Discovery Institute Press). Schon der Titel macht deutlich, dass er seine im Jahr 1985 formulierte Kritik an Evolution auch nach vielen neuen Erkenntnissen und vielen neuen Fossilfunden bestätigt sieht. Die Hauptthese lautete damals: Die Natur ist im Wesentlichen diskontinuierlich; es gibt systematische Sprünge. Höhere Taxa* und Taxa-definierende homologe* Merkmale sind deutlich abgrenzbar.
*Begriffe:
- Taxa (= systematische Einheiten)
- homolog (= bauplangleich)
Wenn Denton Recht hat, ist das ein fundamentales Problem für (neo-)darwinistische Vorstellungen von einer graduellen Evolution, die durch das Wechselspiel von zukunftsblinden Mutationen und gegenwartsbezogener natürlicher Auslese vorangetrieben wird – auch wenn Denton das Evolutionsparadigma als Rahmen akzeptiert.
Strukturalismus und Funktionalismus. Die Problematik für Evolution wird nach Denton durch die Gegenüberstellung von Strukturalismus und Funktionalismus deutlich. Nach dem Strukturalismus sind wesentliche Teile des Bauplans von Lebewesen (Homologien, höhere Taxa definierende Merkmale) durch grundlegende interne Zwänge oder durch physikalische Faktoren bestimmt. Die Strukturen sind primär und ermöglichen Funktionen.
Dagegen sind nach dem Funktionalismus die Funktionen der Organe primär. Diese Sicht ist für die (neo-)darwinistische Theorie grundlegend: Die Organe seien während der Evolution aufgrund funktioneller Notwendigkeiten durch kumulative Selektion und Anpassung entstanden – in vielen kleinen Schritten, wie Darwin betonte. Nicht physikalische Gesetzmäßigkeiten und innere Zwänge sind entscheidend, sondern von Beginn an und jederzeit Anpassung an die Umwelt und die Funktion des jeweiligen Organs. Der Gegensatz beider Anschauungen kann in einer Frage so gefasst werden: Bestimmt die Struktur die Funktion oder bestimmt die Funktion die Struktur?
Dentons Buch ist ein Plädoyer für den Strukturalismus und somit eine Kritik des Darwinismus und der darauf aufbauenden neueren Synthesen. Das zeigt er anhand einer großen Anzahl verschiedenster Beispiele und Phänomene und auch durch Zitate zeitgenössischer Wissenschaftler. Denton legt dar, dass die wesentlichen Bauplan-Merkmale weder durch heutige funktionelle Erfordernisse verstanden werden können noch durch die Annahme, dass ursprünglich (bei ihrer Entstehung) ihre Funktion entscheidend war. Anpassungsvorgänge und Selektion spielen nur eine untergeordnete Rolle bei der mikroevolutiven Feinabstimmung von Organen auf unterschiedliche Umweltbedingungen
Das Auftreten von Neuheiten sei also nicht primär durch funktionelle Notwendigkeit begründet, wie von (neo-)darwinistischen Ansätzen angenommen, sondern durch interne und naturgesetzliche Zwänge bedingt; nur deren spezielle Anpassungen könnten darwinistisch erklärt werden.
Interessant sind Dentons Ausführungen zur Wissenschaftsgeschichte. Er zeigt, dass Charles Darwin und seine Nachfolger in der Sache zu Unrecht als Sieger im Theorienwettstreit zur Entstehung der Baupläne erklärt wurden. Die nach Darwin übliche Darstellung, die Biologen vor Darwin hätten ihre Überzeugungen auf ihren metaphysischen Glauben gegründet, stelle die Realität auf den Kopf und sei ein Mythos, den die Neodarwinisten geschaffen hätten. Denn in Wirklichkeit hätten die Biologen vor Darwin ihre Sicht von Typen des Lebens als unveränderliche Bestandteile der Weltordnung gerade nicht aus metaphysischen Vorgaben abgeleitet, sondern aus empirischen Befunden.
Kritik. Dentons Grundthese lautet: Das Leben und seine Bauelemente sind integrale Bestandteile der Natur bzw. der kosmischen Ordnung und entstehen wie die Atome oder Kristalle letztlich aufgrund der Eigenschaften der Materie mit Notwendigkeit. Diese Grundthese versucht er mit einer Analogie aus dem anorganischen Bereich zu untermauern: den Formen, die Wasser bilden kann. Doch eine solche Argumentation ist höchst fragwürdig. Denn den anorganischen Beispielen fehlt eine entscheidende Eigenschaft, die die Strukturen von Lebewesen aufweisen: funktionale Komplexität. Lebewesen machen sich aktiv die Naturgesetzmäßigkeiten zunutze; sie sind mit Möglichkeiten ausgestattet, dies tun zu können. Ihre Organe und ihr ganzer „Betrieb“ arbeiten den natürlichen Zerfallstendenzen entgegen. Im nicht-lebenden Bereich der Schöpfung fehlt genau das.
So bleibt vage und unklar, inwiefern die Grundstrukturen der Lebewesen eine Folge von Gesetzmäßigkeiten und internen Zwängen seien, sprich wie diese die Strukturen der Lebewesen hervorbringen. Denton zeigt mit vielerlei Argumenten und gut begründet, dass kleinschrittige Anpassungen keinen Weg zu den Basisstrukturen des Lebens ebnen, da zahlreiche morphologische, physiologische und genetische Änderungen en bloc neu auftreten und aufeinander abgestimmt sein müssen, und dass somit die (neo-)darwinistische Erklärung nicht zielführend ist. Aber die strukturalistische Alternative bleibt im Ungefähren. Gelegentlich spricht Denton von „Vor-Arrangements“ oder Prädispositionen. Nur wenn die Natur speziell bzw. gezielt vor-arrangiert war für die Verwirklichung von Neuheiten, sei deren sprunghafte Entstehung denkbar. Aber woher kommt dann dieses Vor-Arrangement?
Dentons Kritik am funktionalistischen Ansatz erscheint zwar gut begründet, doch kann er seinen strukturalistischen Ansatz genau so wenig empirisch begründen. Seine Kritik spielt jedoch dem Schöpfungsdenken in die Karten und die von ihm vorgetragenen Befunde können als klare Hinweise auf einen Schöpfer gewertet werden, so wie es die Typologen (wie Cuvier, Owen und von Baer) im 18. und 19. Jahrhundert vertreten haben.
Eine ausführliche Buchbesprechung ist unter http://www.wort-und-wissen.de/info/rezens/b56.html frei verfügbar.
Autor dieser News: Reinhard Junker
© 2017, http://www.genesisnet.info/schoepfung_evolution/n249.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
01.04.14 25 Jahre Evolution in vitro
Gastbeitrag von Daniel Vedder
Ein Langzeit-Evolutionsexperiment mit Bakterien erreicht demnächst 60.000 Generationen. In den letzten Jahren wurden bereits interessante Ergebnisse publiziert, etwa zum Auftauchen aerober Citratnutzung in E. coli oder über die Rolle von Hypermutatoren. Nun ist eine neue Studie erschienen, die nahelegt, dass die evolutionäre Adaptation auch bei gleichbleibenden Bedingungen nicht aufhört. Dass neue Strukturen evolutionär entstehen können, wurde bisher jedoch nicht gezeigt.
Das Langzeit-Evolutionsexperiment. Eines der größten Hindernisse der experimentellen Evolutionsbiologie sind die sehr langen Zeitspannen, mit denen gerechnet werden muss. Zwar lassen sich im Freiland Populationen unter mehr oder minder natürlichen Bedingungen beobachten, doch immer nur für relativ wenige Generationen. Die Generationszeit der Studienorganismen erweist sich in der Praxis als begrenzender Faktor, weswegen nur wenige Experimente länger laufen als 100 oder sogar 1000 Generationen. Ein Freiland-Experiment mit Anolis-Eidechsen etwa umfasste 15 Generationen in ebenso vielen Jahren (vgl. Vedder 2012). Wenn von einer Erdgeschichte von mehreren Milliarden Jahren ausgegangen wird, leidet natürlich die Aussagekraft der Ergebnisse, die aus Experimenten mit so wenigen Generationen gewonnen werden.
Um diese Problematik zu umgehen, werden in Evolutionsexperimenten bevorzugt Modellorganismen mit einer möglichst kurzen Generationsdauer eingesetzt, wie die Taufliege Drosophila melanogaster oder der Fadenwurm Caenorhabditis elegans, deren Reproduktionszeit in Tagen oder Wochen statt in Monaten oder Jahren gemessen wird. Dadurch lassen sich in relativ kurzer Zeit viele Generationen beobachten und so aussagekräftigere Ergebnisse erzielen.
Ein anderer beliebter Modellorganismus für solche Experimente ist das Darmbakterium Escherichia coli. Es bietet zwar als Einzeller nur begrenzte Forschungsmöglichkeiten, durchläuft dafür aber an einem einzigen Tag sechs bis sieben Generationen. Aus diesem Grund entschied sich der amerikanische Evolutionsbiologe Richard Lenski, mit E. coli als Studienorganismus zu arbeiten, als er sein Langzeit-Evolutionsexperiment (Long-Term Evolution Experiment, LTEE) startete. Für seine Versuche startete er mit 12 verschiedenen Bakterienpopulationen („Stämme“) in einem Glucose-Nährmedium. Jeden Tag werden aus jedem Stamm einige Bakterien nach dem Zufallsprinzip entnommen und in frisches Medium gegeben, in welchem sie sich weiterentwickeln können. Regelmäßig eingefrorene Proben ergeben eine Art „Fossilbericht“, der benutzt werden kann, um Bakterien unterschiedlicher Generationen miteinander bezüglich evolutionärer Fitness oder sonstiger Eigenschaften zu vergleichen und ggf. auch ausgewählte Etappen zu wiederholen.
Was 1988 als kleines Projekt begann, wurde bald weithin bekannt und erregte immer mehr Aufsehen. Anlässlich des 25-jährigen Jubiläums publizierte die Wissenschaftszeitschrift „Science“ letztes Jahr sogar einen eigenen Bericht darüber (Pennisi 2013). So sind die bis jetzt erreichten ca. 60.000 Generationen rein rechnerisch vergleichbar mit einer Million Jahren Evolution des Menschen, womit es das derzeit mit Abstand am längsten laufende Evolutionsexperiment ist.
Das LTEE hat bisher einige interessante Ergebnisse hervorgebracht: Beispielsweise sind sechs der Stämme zu so genannten Hypermutatoren geworden, d. h. ihre Mutationsrate ist um mehrere Größenordnungen gestiegen. Unerwartet war, dass sich die Stämme trotzdem insofern ziemlich parallel entwickelten, als der Fitnessgraph jedes Stammes dieselbe Form aufwies. Das bekannteste und bedeutendste Ergebnis ist jedoch das Auftauchen eines Citrattransports in E. coli, das für viel Furore sorgte. Die Charakterisierung von E. coli zeigt, dass die Mikroben unter aeroben Bedingungen Citrat nicht in die Zelle transportieren können, und daher unter solchen Bedingungen das Citrat trotz des verfügbaren Zitronensäurezyklus nicht als Kohlenstoffquelle nutzen kann. Blount et al. (2008) hatten berichtet, dass in einem der 12 E. coli-Stämme diese Funktion über mehrere Mutationen erlangt worden ist, dies war als „evolutionäre Neuheit“ beschrieben worden. Bei genaueren Untersuchungen stellte sich jedoch heraus, dass die vermeintliche „key innovation“ lediglich auf Veränderungen in der Genregulation zurückzuführen ist, also keineswegs die Entstehung einer neuen Funktion vorliegt (vgl. Binder 2012).
Zwischendurch experimentierte Lenski auch mit digitalen Organismen (kleine selbst-replizierende Computerprogamme), die eine noch viel kürzere Generationszeit aufweisen und außerdem einfacher zu handhaben und untersuchen sind als Bakterien. Die Relevanz dieser Ergebnisse für reale biologische Systeme ist aber zweifelhaft (vgl. Bertsch und Waldminghaus 2005).
Unendliche Evolution? Neueste Ergebnisse aus dem Lenski-Labor beziehen sich auf den Gesamt-Fitnessgewinn der Populationen, der über 50.000 Generationen des Experiments hinweg gemessen wurde (Wiser et al. 2013). Im Schnitt waren die Bakterien in dem verwendeten Nährmedium 1,7-mal fitter als die ursprüngliche Gründerpopulation, gemessen an ihrer relativen Wachstumsrate. In den frühen Phasen des LTEE stieg die Fitness am schnellsten, danach flachte die Kurve ab, stieg aber immer weiter. Nach 10.000 Generationen hatten die Forscher vorausgesagt, dass es irgendwann keine vorteilhaften Mutationen für die gleichbleibenden Bedingungen geben und die Fitness somit nicht weiter gesteigert würde (Lenski & Travisano 1994). Es gibt jedoch immer noch keine Anzeichen für ein Fitnessplateau. Im Gegenteil, mathematische Beschreibungen der Fitnesskurve über die Zeit legen ein Modell nahe, dessen Steigung nie Null wird, d. h. dass die Fitness also immer größer wird. Interessanterweise haben die oben erwähnten Hypermutatoren fast durchgehend eine höhere Fitness als Stämme mit normalen Mutationsraten.
Diskussion. Dass die Fitness scheinbar unendlich steigt, ist ein unerwartetes Ergebnis. Nach dem gängigen Modell der Fitnesslandschaft entwickeln sich Populationen unter gleichbleibenden Bedingungen in Richtung einer „Bergspitze“, eines Maximums. Ist die Spitze der Landschaft erreicht, kann die Anpassung mangels Möglichkeiten nicht weiter verbessert werden. Natürlich ist es noch möglich, dass das LTEE diesen Zustand trotzdem irgendwann erreichen wird. Pennisi zitiert jedoch Lenski mit folgenden Worten: „Das Konzept einer Fitness-Spitze ist schwerer zu greifen als ich erwartet hatte. Ich halte es für gut möglich, dass die Fitness sich noch eine Million Jahre lang weiter steigern wird“ (Pennisi 2013, übersetzt vom Autor). Auch aus der Sicht der Schöpfungsforschung ist das durchaus überraschend. Lönnigs Regel der rekurrenten Variation besagt, dass einem Grundtyp nur endlich viele verschiedene Mutationen zur Verfügung stehen (vgl. Lönnig 2005). Daher würde man auch hier schlussendlich ein Fitnessmaximum erwarten. Warum dies (noch) nicht eingetroffen ist, bleibt offen. Es muss allerdings auch bedacht werden, dass Fitnesssteigerung nicht gleichbedeutend ist mit der Entwicklung evolutionärer Neuheiten.
Außerdem ist bei der Bewertung der Ergebnisse natürlich zu berücksichtigen, dass dies nur die Daten aus einem einzigen Großexperiment sind. Weitere Ergebnisse könnten das Bild stark verändern. Sollte das aber nicht der Fall sein, müssen einige theoretische Modelle modifiziert oder gar verworfen werden.
Die hervorgehobene Rolle der Hypermutatoren ist auch eine besondere Betrachtung wert. Da die meisten Mutationen der Fitness abträglich sind, war man früher davon ausgegangen, dass die natürliche Selektion die Mutationsrate so gering wie möglich halten würde. Daher mussten, als 1997 im LTEE die Hypermutatoren beschrieben wurden, die sich gegen Nicht-Mutatoren durchsetzen konnten, neue evolutionsbiologische Modelle zum Einfluss der Mutationsrate entwickelt werden (Sniegowski et al. 1997). Der Biochemiker Michael Behe macht außerdem darauf aufmerksam, dass einige der Hypermutatoren-Stämme ihren Phänotyp durch die Ausschaltung zweier Enzyme erlangten. Somit wäre diese evolutionär vorteilhafte Entwicklung eine Rückentwicklung – „ein schlechtes Omen für jegliche Theorie der Evolution, die sich allein auf blinde, ungerichtete Prozesse verlässt“, wie er sich ausdrückt (Behe 2013, übersetzt vom Autor). Ob zudem Hypermutatoren makroevolutive Änderungen erleichtern oder ermöglichen, ist offen und durch die bisher vorliegenden Befunde aus dem LTEE nicht entscheidbar.
Das LTEE bleibt auch nach 25 Jahren ein faszinierendes Experiment, das viele wertvolle Daten liefert. Vielleicht führen einige davon dazu, dass momentan anerkannte Hypothesen revidiert werden müssen – das ist der Gang der Wissenschaft. Ob nun die natürliche Selektion tatsächlich unendlich weiterläuft, wie Lenski vermutet, oder ob sie doch in einem Plateau mündet müssen zukünftige Untersuchungen zeigen.
Literatur
Bertsch E & Waldminghaus T (2005) Evolution virtueller Lebewesen. Stud. Integr. J. 12, 34-35. (http://www.wort-und-wissen.de/sij/sij121/sij121-7.html)
Binder H (2012) Von der Citrat-Verwertung zur Entstehung des Auges?, https://www.genesisnet.info/index.php?News=187
Behe M (2013) Lenski’s Long-Term Evolution Experiment: 25 Years and Counting. Evolution News, http://www.evolutionnews.org/2013/11/richard_lenskis079401.html
Blount ZD, Borland CZ & Lenski RE (2008) Historical contingency and the evolution of a key innovation in an experimental population of Escherichia coli. Proc. Natl. Acad. Sci. 105, 7899-7906
Lenski RE (2011) Evolution in Action – a 50,000 Generation Salute to Charles Darwin. Microbe 6, 30-33.
Lenski RE & Travisano M (1994) Dynamics of adaptation and diversification: a 10,000-generation experiment with bacterial populations. Proc. Natl. Acad. Sci. 91, 6808-6814.
Lönnig WE (2005) Mutation Breeding, Evolution and the Law of Recurrent Variation. Recent Research Developments in Genetics and Breeding 2, 45-70.
Pennisi E (2013) The Man Who Bottled Evolution. Science 342, 790-793.
Sniegowski PD, Gerrish PJ & Lenski RE (1997) Evolution of high mutation rates in experimental populations of E. coli“ Nature 387, 703-705.
Vedder D (2012) Gründereffekt bei Eidechsen: ein Freilandexperiment auf Inseln. Stud. Integr. J. 19, 107-109.
Wiser MJ, Ribeck N & Lenski RE (2013) „Long-term dynamics of adaption in asexual populations. Science 342, 1364-1367.
Autor dieser News: Daniel Vedder
© 2014, http://www.genesisnet.info/schoepfung_evolution/n208.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
10.05.12 Anpassung durch Verluste
Hughes (2011) stellt ein Modell vor, das schnelle Anpassung und Evolution auf der Basis von Plastizität (umweltbedingte Änderungsmöglichkeiten), ausmerzender („purifying“) Selektion und Gendrift verständlich machen soll („PRM-Modell“, „plasticity-relaxation-mutation mechanism“). Dieses Modell könnte manche Befunde besser erklären als die klassische Synthetische Evolutionstheorie, eignet sich aber nicht als Erklärung der Entstehung evolutiver Neuheiten (Makroevolution; vgl. Mikro- und Makroevolution, https://www.genesisnet.info/index.php?Artikel=41223&Sprache=de&l=1).
Austin L. Hughes von der University of South Carolina schlägt in einem kürzlich veröffentlichten Überblicksartikel einen Evolutionsmechanismus ohne positive Selektion vor. Unter positiver Selektion versteht Hughes die Fixierung eines neuen oder bisher seltenen Allels (= Genvariante) durch die Wirkung von Selektion. Im ersten Teil seines Artikels weist er darauf hin, dass es nur wenige gut dokumentierte Beispiele für die Leistungsfähigkeit positiver Selektion gebe; es sei auch methodisch schwierig, positive Selektion auf molekularer Ebene nachzuweisen.
Anstelle positiver Selektion sollen phänotypische Plastizität (durch Umweltbedingungen ausgelöste Änderungen) und Änderungen in der Wirkung der eliminativen (= ausmerzenden) Selektion sowie die Gendrift (= zufällige Änderungen von Allelhäufigkeiten ohne Selektion) die hauptsächlichen Faktoren der phänotypischen Evolution sein. Hughes bezeichnet den vorgeschlagenen Mechanismus als PRM-Mechanismus: „plasticity-relaxation-mutation mechanism“. Dahinter verbirgt sich folgendes Konzept:
– Alle Lebewesen sind „plastisch“, d. h. sie können je nach Umwelteinflüssen Merkmale unterschiedlich ausprägen.
– Lebt eine Art dauerhaft in einer relativ konstanten Umgebung, werden bestimmte plastische Fähigkeiten nicht genutzt, weil sie nicht (mehr) benötigt werden, d. h. die ausgeprägte Plastizität wird geringer.
– Die Folge ist, dass durch Mutation auftretende Verluste eines Entwicklungswegs nicht mehr durch Selektion ausgemerzt werden. Denn die Phänotypen (= äußere Gestalt), die der eliminativen Selektion („purifying selection“) zum Opfer fallen würden, werden ohnehin nicht ausgeprägt. Im Laufe der Zeit gehen also aufgrund der Verlustmutationen nicht mehr benötigte optionale Fähigkeiten verloren und diese sind dann auch nicht mehr bei Bedarf abrufbar. Diese in der neuen Umwelt neutralen Mutationen können durch Gendrift fixiert werden.
Der PRM-Mechanismus könne – so Hughes – leicht Fälle von explosiver adaptiver Radiation erklären, ebenso die zunehmenden Beispiele schneller ökologischer Anpassungen. Hughes nennt einige Hinweise dafür, dass der PRM-Mechanismus verbreitet sei und diskutiert Testmöglichkeiten.
Wird die Entstehung evolutiver Neuheiten erklärt? Diese Frage kann eindeutig verneint werden. Denn der durch das PRM-Modell beschriebene Vorgang ist sowohl genetisch als auch phänotypisch ausschließlich ein Verlustprozess. Die durch den PRM-Mechanismus erreichte Anpassung beruht in doppelter Hinsicht auf einem Verlust: zum einen auf Verlustmutationen (die nicht ausgemerzt werden können, weil die betroffenen ontogenetischen Entwicklungswege nicht mehr genutzt und damit nicht mehr der Selektion ausgesetzt werden) und zum anderen auf den damit einhergehenden Verlust ursprünglich möglicher Entwicklungswege (was die Plastizität und Anpassungsfähigkeit an verschiedene Umweltbedingungen einschränkt). Chevin & Beckerman (2011, 1) kritisieren, dass das PRM-Modell nicht Anpassungen, sondern Spezialisierungen erklären würde; man kann aber wohl sagen, dass es engere Anpassungen erklärt, was auf „Spezialisierung“ hinausläuft. Chevin & Beckerman meinen vermutlich, dass der Ursprung der betreffenden Merkmale nicht erklärt wird, womit sie Recht haben dürften.
Daher muss bezweifelt werden, ob Radiationen größeren Ausmaßes wie etwa die Entstehung der verschiedenen Säugerordnungen durch das PRM-Modell beschrieben werden können. Denn eine solche gewaltige Radiation erfordert die Entstehung von vielen tiefgreifenden Neuheiten, die kaum aus einer ursprünglichen Plastizität eines gemeinsamen Vorfahren hervorgegangen sein können. Denn das würde nach dem PRM-Modell bedeuten, dass der gemeinsame Vorfahr der heutigen Säugetiere Fledermäuse, Wale, Huftiere, Seekühe, Ameisenbären und viele andere Baupläne in sich vereinigt hätte, was nicht glaubhaft ist. Dagegen dürfte das PRM-Modell für Radiationen im Rahmen eines deutlich enger abgesteckten Bauplans passend sein, etwa bei der Entstehung der Vielfalt der Buntbarsche (Cichliden) in den ostafrikanischen Seen. Beispiele von Veränderungen dieser Art bewegen sich im Rahmen dessen, was auch experimentell oder in Freilandstudien beobachtet werden kann. In solchen Fällen nachweislich schneller ökologischer Anpassungen (von denen mittlerweile etliche bekannt sind) erscheint das PRM-Modell gegenüber der synthetischen Evolutionstheorie überlegen. Denn das PRM-Modell kann die Schnelligkeit des Erwerbs der unterschiedlichen Anpassungen besser erklären: Die unterschiedlichen Anpassungen müssen nicht erst in einem langwierigen Prozess des seltenen Auftretens und allmählichen Fixierens von vorteilhaften Mutationen erworben werden, sondern stehen zu Begin bereits zur Verfügung.
Literatur
Chevin LM & Beckerman AP (2011) From adaptation to molecular evolution. Heredity, doi: doi:10.1038/hdy.2011.96.
Hughes AL (2011) Evolution of adaptive phenotypic traits without positive Darwinian selection. Heredity, advance online publication. doi:10.1038/hdy.2011.97.
Autor dieser News: Reinhard Junker
© 2012, http://www.genesisnet.info/schoepfung_evolution/n184.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
23.10.12 Von der Citrat-Verwertung zur Entstehung des Auges?
Anmerkungen zu einem vielbeachteten Experiment und zur Frage der Entstehung evolutionärer Neuheiten
Ein vielbeachtetes Langzeit-Evolutions-Experiment scheint entscheidende Hinweise zu den lange gesuchten Mechanismen der evolutiven Entstehung von Innovationen zu geben. Von einem „Schlag ins Gesicht“ der Evolutionskritiker sprechen zwei Kommentatoren in der Wissenschaftszeitschrift „Nature“. Was die neuen Befunde zeigen und welche Fragen offen bleiben wird nachfolgend kommentiert.
Im Jahr 1988 begann Richard E. Lenski mit seiner Arbeitsgruppe ein bislang einzigartiges und sehr aufwändiges Langzeit-Evolutions-Experiment (LZEE) mit Escherichia coli-Bakterien. Aufgrund der kurzen Generationszeit können unter den gewählten Kulturbedingungen im Durchschnitt ca. 6,6 Generationen pro Tag erzeugt werden. Im Jahr 2010 feierten die Forscher die 50.000 Generation, die sie hintereinander gezüchtet und auf evolutive Veränderungen hin untersucht hatten. Mittlerweile sind 55.000 Generationen erreicht. Nach jeweils 500 Generationen werden Proben tiefgefroren und damit ein umfangreiches Archiv von reaktivierbaren Bakterien angelegt. Damit können bei Bedarf die Abfolge der Änderungen zurückverfolgt und Wiederholungsversuche unternommen werden.
Eine neue Funktion
Im Rahmen dieser Versuchsreihen ist eine unüberschaubar große Zahl von Mutanten aufgetreten. Im Jahr 2008 wurde darunter erstmals eine „neue“ Funktion beschrieben (Blount et al. 2008; vgl. Schilderung bei Binder 2008). Diese Mutation (Cit+) versetzte die Bakterien in die Lage, Zitronensäure (Citrat) als Kohlenstoffquelle zu nutzen – eine neue Fähigkeit, die den Bakterien zuvor fehlte. E. coli verfügte zwar bereits über einen vollständigen Zitronensäurezyklus; die notwendigen Stoffwechselprozesse für die Verwertung von Zitronensäure waren also schon vorhanden. Allerdings konnte das Bakterium Citrat aus dem Nährmedium vorher nur unter sauerstofffreien Bedingungen nutzen. In Gegenwart von Sauerstoff konnte kein Citrat aus der Umgebung in die Zelle transportiert werden. Die Fähigkeit zur Aufnahme von Zitronensäure aus dem Nährmedium trat erst nach etwa 31.500 Generationen auf, obwohl Citrat von Anfang an zur Verfügung stand. Zunächst war nicht klar, welche Änderungen zu dieser Fähigkeit geführt hatten.
Mittlerweile konnten die Forscher mit Hilfe ihres Bakterien-Archivs die zugrundeliegenden Vorgänge genauer untersuchen. Schon 2008 hatten die Autoren Hinweise darauf gehabt, dass eine oder mehrere Mutationen vorausgegangen sein müssen, bevor schließlich die Mutation zum Phänotyp Cit+ möglich war. Im LZEE mit E. coli trat zunächst nach ca. 31.500 Generationen eine Variante mit sehr schwacher Cit+-Funktion auf, die mit Cit–-Varianten konkurrierte, ohne diese jemals komplett zu verdrängen. In einem weiteren Ereignis wurde die Effektivität der Nutzung von Citrat (Cit+-Funktion) verbessert. Diese Vorgänge konnten mittlerweile durch weitere Untersuchungen erhellt werden (Blount et al. 2012).
Über den physiologischen Mechanismus, der die Cit+-Funktion ermöglicht, konnte 2008 nur spekuliert werden. Es blieb offen, ob ein „verborgenes“, durch Mutationen funktionslos gewordenes Transportprotein wieder reaktiviert worden ist (was Blount et al. [2008] aufgrund der langen Nichtbenutzung für unwahrscheinlich halten) oder ob ein funktionsfähiges Transportprotein umgebaut und für den Transport von Zitronensäure durch die Membran nutzbar gemacht wurde (diese Vorstellung favorisieren die Autoren).
Wie stellt sich die Sachlage nun dar?
Die neuen Untersuchungen (Blount et al. 2012) haben ergeben, dass sich zunächst eine genetische Veränderung ereignet haben muss (oder zwei Veränderungen), die die Entstehung der Cit+-Varianten überhaupt erst ermöglichte. Um was für eine Art von „ermöglichender“ Mutation es sich dabei handelt, ist unklar, ebenso ob sie einen Selektionsvorteil brachte und sich deshalb durchsetzen konnte oder ob sie (fast) neutral war und sich zufällig ausbreitete (Gendrift). Ein weiterer Schritt ermöglichte den Bakterienstämmen dann eine sehr geringe Verwertung von Citrat („Verwirklichungs“-Mutation). Über ihn wissen die Forscher Genaueres: Betroffen ist das Gen citT, welches ein Transportprotein kodiert, das Zitrat in die Zelle einschleust. „Ursprünglich lag citT einmal unterhalb des Genorts von citG (einem weiteren für die Zitrat-Verstoffwechselung notwendigen Gen) sowie von rnk, einem Gen mit ganz anderem Aufgabenbereich im Energiestoffwechsel. In sämtlichen Cit+-Zellen fiel Blount und seinen Kollegen nun aber ein charakteristisches Rearrangement dieser Gene ins Auge, das rnk mit citG verschmolz. Das führte nun dazu, dass die Expression von citG und citT unter die Promotorkontrolle von rnk geriet. Die Regulatorsequenz dieses Gens erlaubte daraufhin, dass beide cit-Gene nun auch in Gegenwart von Sauerstoff aktiv werden“ (Hendricksen & Rainey 2012). Es wurde also die Regulation der entsprechenden Gene geändert durch Umordnung eines Genkomplexes. Damit war bereits der entscheidende Schritt zur neuen Funktion geschafft, wenn diese auch sehr schwach ausgeprägt war und den Bakterien vermutlich keine Vorteile brachte. Die weiteren Veränderungen beinhalten dann Optimierungen. Die schwach ausgeprägte Cit+-Funktion wurde in weiteren Schritten dadurch verbessert, dass die Gene in Tandems von zwei bis neun Kopien dupliziert wurden.
Welche Schlüsse können gezogen werden?
Evolution ja … Die immense Zahl an Generationen in den vielen parallel gezüchteten Linien von Bakterien ermöglichte das Studium mehrerer hintereinander erfolgter Mutationen. Keine Frage, in diesem Sinne findet Evolution statt, es treten Änderungen auf, die für die Bakterien unter bestimmten Bedingungen nützlich sind. Mutationen können nachweislich auch zu Umorganisierung von Genen führen. Es ist also möglich, dass die Regulation von Genen geändert wird (was keine neue Erkenntnis ist) und es ist ebenfalls möglich, dass sich mehrere Mutationen sukzessive ansammeln, die in der Summe zu einem veränderten Phänotyp führen. Die Arbeiten in Lenskis Labor dokumentieren solche graduellen Änderungen in mehreren aufeinander folgenden Mutationsschritten. Nach gegenwärtigem Kenntnisstand brachte erst der dritte Schritt dem Organismus überhaupt einen Vorteil (die ersten Schritte waren vermutlich neutral, was aber nicht sicher ist, s. u.) und führte zu einer sprunghaften Änderung des Phänotyps (Nutzung von Citrat als Nährstoff). Vorgänge dieser Art waren von manchen Evolutionskritikern bisher bezweifelt worden.1 Die nachgewiesene Anhäufung mehrerer Mutationen ist ein sehr interessanter Aspekt unter den Ergebnissen von Lenskis Arbeitsgruppe. Derzeit ist im Blick auf die erste „ermöglichenden“ Mutation allerdings wenig bekannt, ebenso ist die Art und der Grund ihrer Fixierung in den Populationen unklar.
… aber von welcher Qualität? In der entsprechenden Nature-Ausgabe kommentieren Hendricksen & Rainey (2012) die neuen Ergebnisse des LZEE. Dabei werfen sie Eingangs die Frage auf, welche kleinen Schritte einem Sprung wie etwa der Erfindung des Konstruktionsprinzips „Auge“ vorausgehen. Damit wird nahegelegt, dass die beiden Probleme – Aktivierung des bereits vorhandenen Citratimports unter veränderten Bedingungen und die Entstehung des Auges – vergleichbar wären. Davon kann aber nicht im Entferntesten die Rede sein. Später sprechen sie von der Entstehung „völlig anderer Phänotypen“, was eine gewaltige Übertreibung darstellt. Lenskis Befunde beleuchten Vorgänge im Zusammenhang mit der Fähigkeit von Organismen, sich veränderten Bedingungen anzupassen. Aber in keiner Weise geben sie einen Hinweis darauf, wie zum Beispiel ein Stoffwechselweg de novo entsteht. Experimentell nachgewiesen sind Vorgänge der Änderung von Regulationsprozessen und in sehr bescheidenem Maße auch die Rekrutierung vorhandener Gene in einem neuen Zusammenhang (vgl. auch Rebeiz et al. 2011). Die nachgewiesenen Veränderungen bleiben jedoch in einem engen Rahmen und jede weitergehende Behauptung erfordert eine eigene Begründung. Der von Hendricksen & Rainey (2012) sogar beschworene „Kampf ‚Wissenschaft gegen Dogma‘“ ist hier ein viel zu starkes und nicht gerechtfertigtes Bild, es bedient nur ein verbreitetes Klischee. Es geht doch nur um die Frage, welche Schlussfolgerungen die empirischen Befunde erlauben und welche nicht.
Was ist eine Neuheit? Auffällig nicht nur bei der Kommentierung ist die Unklarheit, was unter einer „evolutionären Neuheit“ verstanden wird. Der Begriff (oder sinngemäß ähnliche Begriffe wie „key innovation“ – Schlüsselinnovation) wird oft undifferenziert verwendet, d. h. ohne ihn näher zu bestimmen.2 Wenn Änderungen, die keinerlei neue Stoffwechselmoleküle oder gar neue Stoffwechselkaskaden erfordern, zu etwas „völlig Neuem“ hochstilisiert werden, dann erscheint das unangemessen. Von der Änderung von Regulationen, die zum Ausnutzen eines bereits vorhandenen Stoffwechselwegs führen, auf weitergehende und andersartige Änderungen zu schließen, ist nicht zulässig. Hier wird auch außer Acht gelassen, dass es viele verschiedene Grade von Unterschieden biologischer Strukturen, Stoffwechselkomplexen, Signalkaskaden oder molekularen Maschinen gibt. Daher muss jeder Einzelfall gesondert auf seine Aussagekraft im Hinblick auf das evolutionäre Potential natürlicher Mechanismen beurteilt werden.
Die Experimente der Arbeitsgruppe von Lenksi sind spannend und lehrreich, und man kann sicher weitere interessante Ergebnisse erwarten. Eine an den Befunden orientierte Deutung sollte aber die Grenzen des Begründbaren respektieren. Wenn diese Ergebnisse in verschiedenen Ursprungsmodellen gedeutet werden, sollte dies nüchtern und differenziert erfolgen, die dabei vorgenommenen Grenzüberschreitungen sollten dabei deutlich markiert werden.
Anmerkungen
1 Michael Behe, durch sein Buch „Darwin‘s Black Box“ bekannter Evolutionskritiker, hat in seinem Nachfolgewerk „The Edge of Evolution“ (Behe 2007) solche Vorgänge am Beispiel der Entstehung von Frostschutzproteinen dagegen ausdrücklich als plausibel anerkannt.
2 Blount et al. (2012, 513) definieren im Anschluss an Ernst Mayr „evolutionary novelties“ als „qualitatively new traits that open up ecological opportunities and thereby promote diversification“. Was ein „qualitativ neues Merkmal“ ist, müsste nun aber genauer beschrieben werden.
Literatur
Behe M (2007) The Edge of Evolution. The search for the limits of Darwinism. New York.
Binder H (2008) Langzeit-Evolutionsexperiment mit Escherichia coli. Empirischer Befund für neue Funktion durch Mutation? Stud. Int. J. 15, 96-98.
Blount ZD, Borland CZ & Lenski RE (2008) Historical contingency and the evolution of a key innovation in an experimental population of Escherichia coli. Proc. Natl. Acad. Sci. USA 105, 7899-7906.
Blount ZD, Barrick JE, Davidson CJ & Lenski RE (2012) Genomic analysis of a key innovation in an experimental Escherichia coli population. Nature 489, 513-518.
Hendrickson H & Rainey PB (2012) Evolution: How the unicorn got its horn. Nature 489, 504-505. Deutsche Übersetzung unter http://www.spektrum.de/alias/evolution/wie-das-einhorn-zu-seinem-horn-kam/1166511
Rebeiz M, Jikomes N, Kassner VA & Carroll SB (2011) Evolutionary origin of a novel gene expression pattern through co-option of the latent activities of existing regulatory sequences. Proc. Natl. Scad. Sci. 108, 10036-10043.
Autor dieser News: Studiengemeinschaft Wort und Wissen
© 2012, http://www.genesisnet.info/schoepfung_evolution/n187.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
25.06.10 Lag Darwin falsch?
Vorstellung eines unorthodoxen Buches über Evolution
Jerry Fodor und Massimo Piattelli-Palmarini haben Anfang dieses Jahres das Buch „What Darwin got wrong“ veröffentlicht (New York 2010). Darin äußern sie grundlegende Kritik an der auf Darwin zurückgehenden Selektionstheorie. Sie behaupten nicht nur, dass Selektionstheorie darin scheitere, die Entstehung neuer Formen zu erklären, sondern dass es derzeit auch keine alternative Erklärung gebe. Sie sind aber überzeugt, dass es eine rein naturalistische Erklärung gibt. Sie begründen ihre Kritik wie folgt: 1. Vielfältige epigenetische* Prozesse und Kontrollmechanismen beeinflussen das phänotypische** Ergebnis von Mutationen massiv, durch Selektion erfolge nur noch eine Feinabstimmung. 2. Selektion auf ein bestimmtes Merkmal hin („selection for“) sei intentional (erfordere einen zielorientiert handelnden Akteur); diesen gebe es zwar in der Züchtung, nicht aber unter natürlichen Verhältnissen. Daher sei die künstliche Selektion als Vergleich für natürliche Selektion ungeeignet. 3. Evolution sei als historischer Prozess ausgeprägt mehrstufig und nicht gesetzmäßig beschreibbar, sondern eher ein Bündel evolutionärer Szenarien. Diese Kritikpunkte werden nachfolgend erläutert. Die Autoren erhielten teilweise harsche Kritik für ihr Buch, darauf wird abschließend kurz eingegangen.
*epigenetisch (= das Ablesen und Umsetzen des Erbguts beeinflussende)
**phänotypisch (= das äußere Erscheinungsbild betreffend)
Die Autoren1 beginnen überraschenderweise mit dem Hinweis, dass ihr Buch nicht von Gott handle, auch nicht von Intelligent Design und Kreationismus. Damit wollen sie einer Vereinnahmung durch Schöpfungsgläubige vorbeugen, denn ihr Anliegen ist zu zeigen, dass die Theorie der natürlichen Selektion einem verhängnisvollen Irrtum unterliegt. „Heutzutage haben Biologen guten Grund zu glauben, dass Selektion von zufällig erzeugten kleineren Varianten phänotypischer Merkmale zur Erklärung neuer Formen des Lebens bei weitem nicht ausreicht“ (21).2 Das ist keine Kleinigkeit, denn diese Theorie ist ein wesentlicher Teil der kausalen Evolutionstheorien bis heute, wenn nicht ihr Kernstück. Um den Lesern Klarheit über ihre eigene Position zu verschaffen, bekennen sich die beiden Autoren auch noch gleich als Atheisten, die uneingeschränkt naturalistische Erklärungen suchen. Wenn also mit der Selektionstheorie etwas grundlegend verkehrt ist, so muss für die Autoren eine Alternative eine ebenso naturalistische Erklärung bieten, wie es die Selektionstheorie nach deren Befürworter bieten soll. „Es ist unsere Annahme, dass Evolution durch und durch ein mechanischer Prozess ist“ (xiii) – erfreulich, dass sie hier von „Annahme“ sprechen. Sie sind soweit auf Darwins Seite.
Ein weiteres stellen die Autoren schon auf den ersten beiden Seiten klar: Ihr Buch kritisiert – vor allem das, was in ihren Augen falsch ist beim Darwinismus. Nur gegen Ende des Buches sprechen sie eine Alternative an, die nach ihren eigenen Worten „ziemlich vage“ ist: „In der Tat wissen wir nicht sehr gut, wie Evolution funktioniert. Darwin wusste es auch nicht und (soweit wir das sagen können) auch sonst niemand“ (xiv).3 Im letzten Kapitel wiederholen sie diese Einschätzung: „’OK, wenn also Darwin falsch lag, was denkt ihr ist der Mechanismus der Evolution?’ Kurze Antwort: Wir wissen nicht was der Mechanismus der Evolution ist“ (153)4, sie präsentieren dann einige sehr theoretische Überlegungen, in welcher Richtung eine Antwort liegen könnte.
Kurzer Überblick. In Kapitel 1 zeigen die Autoren, inwiefern die Lerntheorie von B. F. Skinners Behaviourismus und der Neodarwinismus ähnlich sind. Beide arbeiten mit einem Modell eines „Generators und Filters“, mit dem die Phänomene erklärt werden sollen; der Generator funktioniert nach dem Zufallsprinzip und der Filter ist exogen (außerhalb des Systems). Im Falle der Evolutionstheorie sind das Mutation und Umweltselektion. Die Selektionstheorie sei aus ähnlichen Gründen wie der Behaviourismus gescheitert.
In den Kapiteln 2-4 schildern die Autoren eine größere Anzahl von Fakten und Mechanismen, die nichts mit Selektion zu tun haben und zeigen, warum der „Darwinismus“ (die Selektionstheorie) nach ihrer Einschätzung nicht stimmen kann. In Kapitel 5 werden Beispiele optimaler Strukturen vorgestellt, die im neodarwinistischen Theorierahmen fremd sind.
Ab Kapitel 6 (Teil II) geht es um die logische und konzeptionelle Basis der Theorie der natürlichen Selektion. Sie geben eine Begründung dafür, dass die Rolle von Zufallsgeneratoren (Mutationen) und Umweltfiltern (Selektion durch Umwelteinflüsse) überschätzt worden sind in Bezug auf ihre Fähigkeit, Phänotypen (das Erscheinungsbild) zu formen. In das Anpassungskonzept sei Zielorientierung eingebaut worden, die es in Wirklichkeit nicht gebe. In Kapitel 8 weisen die Autoren das verbreitet vorgebrachte Argument zurück, es gebe keine naturalistische Alternative zur Selektionstheorie, weshalb sie beibehalten werden müsse. Im Schlusskapitel thematisieren sie den Charakter historischer Erklärungen und wenden dies auf die Selektionstheorie an.
Kritikpunkt interne Entwicklungszwänge. Die Erzeugung von Mutation erfolge zwar zufällig, das gelte aber nicht für die daraus resultierenden Phänotypen. Denn bevor das Ergebnis einer Mutation der Umweltselektion „angeboten“ wird, wirken zahlreiche innere (organismusinterne) Faktoren auf mehreren Ebenen und beeinflussen das phänotypische Ergebnis massiv. Eine Reihe von internen Filtern steht zwischen Mutation und ihrer Auswirkung – das ist das Feld von „Evo-Devo“, der evolutionären Entwicklungsbiologie, die das klassische Bild über evolutionäre Veränderungen „ganz beträchtlich“ verändert habe (27). Die Autoren zeichnen ein Panorama spezifischer Mechanismen, deren Entdeckung die gradualistisch-adaptionistische Selektionstheorie mindestens in einigen Fällen als falsch erweist, weil neue phänotypische Merkmale nicht zufällig erscheinen oder weil die Anpassung an die ökologischen Verhältnisse nur eine sekundäre Rolle in der Fixierung der Phänotypen spielt. Dazu gehören Puffersysteme gegen Mutationen; Robustheit, verborgene Variation, Plastizität, verschiedene Regulationsprozesse bei der Transkription, Fehlerkontrollen, Veränderungen nach der Translation (Übersetzung der Erbinformation in Proteine), die Tätigkeit von Chaperonen (Proteine, die Faltungsfehler anderer Proteine korrigieren), alternatives Spleißen und andere Vorgänge. Daher kommen die Autoren zur Einschätzung: „Wir können natürliche Selektion mit dem Stimmen eines Klaviers vergleichen, nicht mit dem Komponieren von Melodien“ (21).5 Die Autoren weisen auch darauf hin, dass lange Zeiträume der Wirkung der Selektion nicht weiterhelfen (S. 21).
Wichtig ist auch die Entdeckung, dass es der Regelfall ist, dass Gene, Proteine und sogar ganze Signalkaskaden mehrfach genutzt werden. Natürliche Selektion kann daher nicht isolierte Merkmale auslesen, sondern nur miteinander verknüpfte Komplexe von Merkmalen.
Neben diesen Rahmenbedingungen (Entwicklungszwänge, constraints) „von unten“, die auf dem Weg vom Genotyp zum Phänotyp die Zufallsmutationen kräftig filtern, bevor sie der Umweltselektion ausgesetzt werden, gibt es auch noch die Rahmenbedingungen „von oben“: die „Gesetze der Form“. Die Rolle der physikalischen Gesetze, die die Möglichkeiten der Gestaltbildung und der Ausbildung von Strukturen der Lebewesen begrenzen, sei oft zu wenig beachtet worden. Viele Formbildungen können nicht durch biochemische Kaskaden, den genetischen Code, Entwicklungspfade und auch nicht durch Selektion verstanden werden, es gebe vielmehr schwerwiegende geometrische und materielle Zwänge, die auf diese Prozesse einwirken.
Die Autoren schildern in diesem Zusammenhang einige Beispiele von optimal und perfekt ausgebildeten Strukturen in der Natur und kommen zum Schluss, dass solche Fälle von ausgeklügelten Programmen nicht durch Optimierung physico-chemischer Faktoren, aber auch nicht durch gradualistische Anpassung erklärt werden können. In der Zukunft mag eine Erklärung möglich sein, es sei aber fair einzuräumen, dass eine Erklärung gegenwärtig fehlt.
Kritikpunkt Trittbrettfahrer und Zielorientierung. Neben der außerordentlichen Vielfalt und Verflechtung der Vorgänge auf dem Weg vom Genotyp zum Phänotyp weisen die Autoren auf ein weiteres grundlegendes Problem mit der Selektionstheorie hin: das Trittbrettfahren (free-riding). In Anknüpfung an einen berühmten Artikel von Gould & Lewontin (1979) zeigen Fodor und Piattelli-Palmarini, dass man an den Merkmalen der Lebewesen nicht erkennen könne, ob sie durch Auslese oder als Nebenprodukt einer Auslese entstanden sind. Gould und Lewontin hatten das Beispiel von Bögen bei Kathedralen gewählt, um zu illustrieren, wie Anpassungen auch zu Merkmalen führen, die gleichsam Trittbrettfahrer sind – in diesem Beispiel die Spandrillen links und rechts der Bögen. Fodor & Piattelli-Palmarini argumentieren nun: Wenn sich ein Phänotyp aufgrund äußerer Selektionsfaktoren ändert, so kann nicht bestimmt werden, welches der neuen phänotypischen Merkmale (wenn überhaupt eines) eine Anpassung und welches (wenn überhaupt eines) ein Trittbrettfahrer ist. In der Zucht sei dies klar, da es einen Ausleser gebe, den Züchter, der allerdings zielorientiert vorgeht. Diese Zielorientierung kann aber in der natürlichen Evolution nicht vorausgesetzt werden. Dort kann es daher zwar Selektion auch ohne Zielorientierung geben, aber nicht Selektion auf etwas hin („selection-for“). Selektion kann nur den gesamten Organismus auslesen, es kann aber in einem naturalistischen Rahmen nicht bestimmt werden, ob ein bestimmtes Merkmal im komplexen Ganzen Gegenstand der Auslese war. Daher sei die künstliche Selektion kein geeignetes Modell für natürliche Selektion und der Begriff „natürliche Selektion“ keine harmlose Metapher. Vielmehr beruhe der Adaptionismus auf der Analogie zur künstlichen Selektion. Das aber sei im Kern verfehlt, da diese intentional (willentlich gesteuert) ist – Darwins Analogie sei also in fataler Weise verkehrt (S. 100). Die Theorie der natürlichen Auslese könne weder vorhersagen noch erklären, welche Merkmale der Lebewesen einer Population Gegenstand der Auslese sind und welche nur Trittbrettfahrer sind (S. 110, 114). In einem intentionalen Kontext – also bei bewusster und gezielter Auslese wäre dies anders zu bewerten, da hier Ziele verfolgt werden, aber das ist bei einer natürlichen Evolution nicht gegeben.
Daher sei das Beispiel von Bögen und Spandrillen durch Gould und Lewontin schlecht gewählt: Hier könne die Selektion auf etwas hin (die Bögen) leicht von den Trittbrettfahrern (den Spandrillen) unterschieden werden, weil eine geistige Verursachung vorliegt, es also einen intelligenten Urheber gebe (S. 115; vgl. S. 12). Diesen Fehler habe auch Darwin gemacht, als er die künstliche Selektion zum Vorbild der natürlichen Selektion verwendete. Man könne aber nicht mit einem mentalen Prozess beginnen und nachher in der Anwendung auf natürliche Vorgänge einfach von ihm absehen (S. 116). Die Kritik müsse darüber hinaus auf alle evolutionären Erklärungen ausgedehnt werden, die Bezug auf Selektion nehmen.
Kritikpunkt historische Theorien. Die Autoren thematisieren als weiteren Problempunkt die Tatsache, dass die „Geschichte über die Evolution der Phänotypen“ nicht zur Biologie, sondern zur Naturgeschichte gehöre. Jegliche Geschichte aber, auch die Naturgeschichte operiert mit Erklärungen, die nicht dem Newtonschen Paradigma einer einstufigen Theorie („single-level“) entsprechen. Vielmehr sind historische Theorien ausgeprägt mehrstufig („multi-level“) und nehmen Bezug auf sehr viele Ursachen. Evolutionäre Theorien würden – so die Autoren – meistens (aber fälschlicherweise) als einstufig betrachtet (S. xx).
Adaptionistische Erklärungen sind historische Erklärungen, diese bieten plausible Erzählungen, eine kausale Kette von Ereignissen, die zum Ereignis führten, das erklärt werden soll. Nomologische Erklärungen handeln dagegen von (metaphysisch notwendigen) Beziehungen von Eigenschaften, während historische Erzählungen von (kausalen) Beziehungen zwischen Ereignissen handeln (132). Historische Erklärungen sind für die Autoren „perfectly OK“, aber sie subsummieren Ereignisse nicht unter Gesetze und unterstützen daher nicht „counterfactuals“, d. h. können keine Antwort auf die Frage geben, was passiert wäre, wenn bestimmte Dinge (das Antezedens, die Ausgangsbedingungen) anders gewesen wären. Die Folgen alternativer Ausgangsbedingungen müssten aber untersucht werden können, um das Trittbrettfahrerproblem zu lösen (etwa: was wäre passiert, wenn die Umstände anders gewesen wären). „Wenn Anpassungstheorien aber eher historisch als gesetzmäßig sind, wird dadurch erklärt, weshalb das Trittbrettfahrerproblem innerhalb des Rahmenwerk des Adaptionismus nicht gelöst werden kann“ (133). Die Autoren stellen fest: „Naturgeschichte ist keine Evolutionstheorie, sie ist ein Bündel evolutionärer Szenarien“ (159).6
Abschließende Bemerkungen
Die beiden Autoren bringen für den Mainstream in der Evolutionsforschung ohne Zweifel eine unpopuläre Botschaft und sie wagen es, von einem zentralen ungelösten Problem zu sprechen, ohne selber eine Lösung zu präsentieren („we don’t know what the mechanism of evolution is“, S. 153). Es überrascht daher nicht, dass sie mit Kritik konfrontiert werden, die wenig sachorientiert ist. So wird beispielsweise unterstellt, sie hätten keinerlei Interesse an den Fakten (das ist pure Diffamierung und wird durch die Ausführungen in ihrem Buch widerlegt), oder man mutmaßt irgendwelche wahren Gründe7 ihrer Kritik, statt auf ihre Argumente einzugehen. Pigliucci (2010) gibt die Argumentation der Autoren ungenügend und verzerrt in seiner Rezension in Nature wieder und baut schon in der Überschrift einen Strohmann: „Attacke auf Evolution“. Seine Rezension verfehlt folgerichtig die Argumentation von Fodor & Piattelli-Palmarini. Ebenso stützt Coyne (2010) seine Kritik an den beiden Autoren auf Auslassungen und Verzerrungen und baut darauf persönliche Diffamierungen auf; man vermisst bei ihm den ernsthaften Versuch, die kritisierten Autoren zu verstehen. David Sloan Wilson bezeichnet Fodor gar als „säkularen Kreationisten“ (in Mazur 2009, 38; online unter http://www.suzanmazur.com/?p=20). Dies sind allzu bekannte und leider verbreitete Mittel in Kontroversen, mit denen die Kontrahenten bzw. ihre Position bekämpft werden. Umso erstaunlicher ist es, dass diese Mittel angewendet werden, obwohl die Autoren sich ausdrücklich und ohne einen Hauch eines Zweifels zum Evolutionsparadigma und einem naturalistischen Weltbild bekennen. Aber selbst unter diesen Vorzeichen scheint eine freie, sachorientierte Diskussion manchen Zeitgenossen nicht zu passen.
Die Autoren selber schreiben (S. xx), dass einige ihrer Kollegen ihnen geraten hätten, ihre Kritik Darwins nicht zu veröffentlichen. Sie hätten es dennoch gemacht mit dem Argument, sie würden sich sonst mit den „Mächten der Finsternis“ zusammentun, deren Ziel es sei, Wissenschaft in Verruf zu bringen. In der Tat: Wenn zentrale Konzepte von Evolutionstheorien nicht öffentlich kritisch diskutiert werden dürften, wäre das an diesen Stellen das Ende der Wissenschaft.
Leitet „What Darwin got wrong“ also Wasser auf die Mühlen des Kreationismus und der Intelligent-Design-Bewegung? Diese von manchen geäußerte Befürchtung ist teilweise vielleicht berechtigt, denn Fodor und Piattelli-Palmarini thematisieren einen wichtigen Aspekt des Design-Ansatzes: Ist ein wirklich durch und durch natürlicher ateleologischer (nicht zielgerichteter) Evolutionsmechanismus bekannt? Und sie verneinen eindeutig, geben aber ebenso eindeutig der Überzeugung Ausdruck, dass es doch eine (wenn auch noch unbekannte) positive Antwort auf diese Frage gibt. Daher ist es nicht angebracht, die Autoren für Überzeugungen zu vereinnahmen, die sie nicht teilen oder sogar ablehnen. Es ist den Autoren hoch anzurechnen, dass sie sich keinen Maulkorb haben verpassen lassen.
Literatur
Coyne J (2010) The improbability pump. http://www.thenation.com/article/improbability-pump.
Gould SJ & Lewontin R (1979) The spandrels of San Marco and the Panglossian paradigm: a critique of the adaptionist programme. Proc. R. Soc. Lond. B 205, 581-198.
Mazur S (2009) The Altenberg 16. An exposé of the evolution industry. Wellington New Zealand.
Pigliucci M (2010) A misguided attack on evolution. Nature 464, 353-354.
Anmerkungen
1 Jerry Fodor ist Professor für Philosophie und Kognitionswissenschaften, Massimo Piattelli-Palmarini arbeitete zunächst als Biophysiker und Molekularbiologe und ist heute Professor für Kognitionswissenschaften.
2 „These days biologists have good reasons to believe that selection among randomly generated minor variants of phenotypic traits falls radically short of explaining the appearance of new forms of life.“ Vgl. auch: Die Theorie der natürlichen Selektion sei „irredeemably flawed“ (1) „Whatever NS [natural selection] is, it cannot be the mechanism that generates the historical taxonomy of species“ (2). „We will argue that, in fact, ET can offer no remotely plausible account how filtering by natural selection might work“ (16).
3 „In fact, we don’t know very well how evolution works. Nor did Darwin, and nor (as far as we can tell) does anybody else.“
4 „’OK, so if Darwin got wrong, what do you guys think is the mechanism of evolution?’ Short answer: we don’t know what the mechanism of evolution is.“
5 „We think of natural selection as tuning the piano, not as composing the melodies.“
6 „Natural history isn’t a theory of evolution; it’s a bundle of evolutionary scenarios.“
7 http://chronicle.com/article/What-Darwins-Doubters-Get/64457/; siehe auch http://www.boston.com/ae/books/articles/2010/02/14/new_critique_intends_to_rebut_darwins_ideas/ und http://www.rsng.de/aktuelles/120210-neuer-angriff-auf-den-darwinismus.php (wo einige gesammelte Reaktionen zu finden sind)
Autor dieser News: Reinhard Junker
© 2010, http://www.genesisnet.info/schoepfung_evolution/n153.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
26.06.08 „Gott als Designer?“
Bericht und Kommentare zu einer Tagung der Evangelischen Akademie Badens
Unter dem Thema „Gott als Designer? Theologie und Naturwissenschaft im Gespräch“ fand vom 6.-8. Juni 2008 an der Evangelischen Akademie Baden in Bad Herrenalb (Nordschwarzwald) eine Tagung über Schöpfung und Evolution statt. Das Thema wurde von sechs Referenten aus biologischer, wissenschaftstheoretischer, theologischer und religionspädagogischer Sicht beleuchtet. Als einziger Evolutionskritiker war auch Reinhard Junker von der Studiengemeinschaft Wort und Wissen mit von der Partie. Folgende Vorträge wurden in dieser Reihenfolge gehalten (mit anschließender meist halbstündiger Diskussion):
Prof. Dr. Thomas Junker: Was ist Evolution? Die Darwinsche Revolution und die moderne Biologie
Dr. Reinhard Junker: Evolution und Schöpfung. Theologische Motive und naturwissenschaftliche Aspekte des Kreationismus
Dr. Hansjörg Hemminger: Mit der Bibel gegen die Evolution? Evolution, Intelligent Design und Kreationismus
Prof. Dr. Jürgen Audretsch: Reichweite und Grenzen naturwissenschaftlicher Aussagen
Prof. Dr. Manfred Oeming: Ist die Bibel ein Tatsachenbericht? Lesehilfen zur Lektüre der Schöpfungsgeschichte
Prof. Dr. Astrid Dinter: Evolutionstheorie gegen Schöpfungstheologie? Eine Auseinandersetzung mit dem Buch „Gotteswahn“ von Richard Dawkins
Es waren gut 100 Teilnehmer anwesend, viele davon gehörten zum Stammpublikum der Evangelischen Akademie, aber es waren auch andere Gäste dabei; die Tagung war besser besucht als vom Veranstalter erwartet.
Nachfolgender Bericht des Mitreferenten Reinhard Junker soll keinen vollständigen Überblick über alle Themen und Thesen geben, sondern greift einige Punkte heraus, die aus der Sicht von Wort und Wissen von besonderer Relevanz oder von besonderem Interesse sind.
Der Biologe Thomas Junker präsentierte einige Inhalte der Evolutionstheorie, vor allem zu den Evolutionsmechanismen: Variation, Selektion und Zeit reichen aus, um die Vielfalt des Lebens hervorzubringen. Er baute darauf einen rigorosen Biologismus auf, so dass es nur folgerichtig war, dass er keinerlei Vereinbarungsmöglichkeiten von Evolution und einem wie auch immer gearteten Schöpfungsglauben sah. Die Evolution könne alles erklären, auch den Sinn des Lebens, der darin bestehe, dass die biologischen Maschinen (auch wir Menschen) ihre Gene verbreiten. Dazu gab es in einem solchen Rahmen verständlicherweise viel Kontra. Ein Teilnehmer meinte, er hätte zwar vier Kinder, aber keine Enkel; ob er nun den Lebenssinn verfehlt habe. – Nein, so die Antwort, denn die Evolution habe dafür gesorgt, dass es einem gut gehen solle, denn das sei eine grundsätzliche Voraussetzung dafür, dass man seine Gene gut weitergeben könne. Damit sei auch ein „hedonistischer Sinn des Lebens biologisch begründet“. Vom Publikum wurde auch kritisch bemerkt, dass Thomas Junker dauernd teleologische (= zielgerichtete) Begriffe für Evolutionsvorgänge verwendet – schließlich kennt Evolution kein Ziel. Antwort: Das sei abkürzende Redeweise. Es waren z. B. folgende Begriffe im Zusammenhang mit Evolution verwendet worden: „Programmierung“, „Zwecke“, „um … zu“-Formulierungen, „egoistische Gene“, „erfinden“, „Experiment“ der Evolution, „Architekt“, „Konstrukteur“. Ich konnte in meinem darauf folgenden Vortrag darauf hinweisen, dass Formulierungen, die ohne teleologische Elemente in den Begriffen auskommen, in vielen Fällen in der Biologie noch gar nicht gefunden worden seien.
Bemerkenswert war noch der Versuch des Referenten, „Evolution – ein kritisches Lehrbuch“ zu diskreditieren; und zwar wie folgt: In dessen Vorwort steht: „Deutungsprobleme und offene Fragen von Evolutionstheorien … haben nach unserer Auffassung ein so großes Gewicht, dass Makroevolution als nicht mehr hinterfragbare Leitvorstellung ernsthaft geprüft und nicht als Tatsache vorausgesetzt werden sollte.“ Thomas Junker nannte es „bodenlosen Unsinn“ zu behaupten, dass Evolution als Tatsache vorausgesetzt werde; das stimme nicht. Man würde ständig Überprüfungen vornehmen. Einige Minuten später betonte er dann sehr stark, dass Evolution eine Tatsache sei – so sicher wie Tatsachen nur sein könnten. Damit hatte er sich selbst widersprochen. Ich konnte das in meinem Vortrag klären und wies auf die Wendung „als nicht mehr hinterfragbare Leitvorstellung“ hin. Außerdem warf er Evolutionskritikern vor, mit den Begriffen „Mikroevolution“ und „Makroevolution“ Verwirrung stiften zu wollen. Tatsächlich trifft aber das Gegenteil zu. Wir bemühen uns um saubere Begriffsklärungen als Basis für eine fruchtbare Diskussion. Die Grundtypenbiologie bezeichnete Thomas Junker als „unkritischen naivsten Wunderglaube“, das sei „Kindergarten“.
Der Hauptteil des Vortrags des Weltanschauungsbeauftragten und Biologen Hansjörg Hemminger bestand aus einem durchaus interessanten und kurzweilig vorgetragenen historischen Rückblick zur Geschichte des Kreationismus und der Intelligent Design-Bewegung. Dessen Ausführlichkeit ließ dann aber kaum Zeit für eine nennenswerte substantielle Auseinandersetzung, weder bezüglich naturwissenschaftlicher noch theologischer Argumente. Die Kritik am Kreationismus erschöpfte sich im Wesentlichen in der Aussage, dass der Kreationismus als Naturwissenschaft in einem Vergleich mit der Evolutionstheorie überall schlechter abschneide; der Kreationismus stelle sich einer Hypothesenkonkurrenz nicht, in den meisten Feldern sei er nicht einmal als Konkurrent angetreten. Hemminger ging kurz auf den „Intelligent Design“-Ansatz (ID) ein, der meines Erachtens unausgewogen dargestellt wurde. Man konnte seinem Vortrag manches darüber entnehmen, was er nicht vertritt, weniger aber, wofür er theologisch steht. Als Motive des Kreationismus, die in dessen Geschichte vorherrschend waren, nannte er die Geltung der Heiligen Schrift (was er als „Bibelfundamentalismus“ bezeichnete), die Rettung des christlichen Menschenbildes (was Folgen für Ethik und Menschenwürde habe) sowie neuerdings vor allem in der ID-Bewegung einen Gottesbeweis auf dem Wege naturwissenschaftlicher Argumentation. Hier überraschte mich sein Optimismus bezüglich der Leistungskraft der Evolutionsmechanismen: Die Entstehung sog. „komplexer spezifizierter Information“ (dazu gehören z. B. biochemische Maschinen wie der Bakterien-Rotationsmotor) sei durch Selbstorganisation problemlos möglich.
Der Physiker Jürgen Audretsch unterschied zwischen starken und schwachen ID-Versionen. Das erscheint mir als eine sehr wichtige Unterscheidung. Nach der starken Fassung könne man – vereinfacht gesagt – einen Designer aus der Natur beweisen, die schwache Fassung dagegen versteht Design-Indizien nur als Veranschaulichung bei Vorgabe eines Designers (gedrehtes Design-Argument). Naturwissenschaft sei Laborwissenschaft (plus Freilandbeobachtungen) und könne ihre Aussagen nur unter Vorgabe von Voraussetzungen machen (was Thomas Junker in der Diskussion zu seinem Vortrag implizit bestritt, denn er meinte, man ginge streng induktiv vor, „da kann Popper sagen was er will“). Begründungsketten müssten in der Naturwissenschaft lückenlos sein, es gebe Gültigkeitsbereiche, aber auch Grenzen der Naturwissenschaft. Dagegen sei Rekonstruktion der Naturgeschichte nicht Teil der Naturwissenschaft, sondern nur Anwendungsgebiet der Naturwissenschaft. Geschichte ist Rekonstruktion bzw. Konstruktion der Vergangenheit, ein Blick zurück auf der Grundlage der vorliegenden Beobachtungsdaten mit Hilfe der naturwissenschaftlichen Methoden und dem naturwissenschaftlichen Wissen der Zeit. Dazu seien die Kriterien der Geschichtswissenschaft anzuwenden. Er stimmte einem Diskussionsbeitrag aus dem Publikum zu, dass es verschiedene Rekonstruktionen geben könne, die miteinander konkurrieren.
Begründungslücken könne es durchaus geben. Audretsch brachte aus der Kosmologie das Beispiel der Standardtheorie, die solche Lücken gehabt habe, die durch die Inflationstheorie zwar gefüllt worden seien, wodurch sich die Lücken aber nur verschoben hätten.
„No-go-Aussagen“ (Unmöglichkeitssätze) (was ja ID auch macht) gebe in großer Zahl in der Wissenschaft, sie seien aber nur unter der Angabe von Voraussetzungen möglich.
Der Alttestamentler Manfred Oeming legte einige Schöpfungstexte aus, sehr ausführlich Gen 1 und Gen 2, sowie Texte aus Jesaja, Hiob und den Psalmen. Seiner textgetreuen Auslegung von Gen 1 konnte ich praktisch uneingeschränkt zustimmen; dagegen fast überhaupt nicht seinen Ausführungen zu Genesis 2. Gott werde dort als Experimentator gesehen, dem auch mal etwas missglücke, so beim Versuch, unter den Tieren einen Partner für den Menschen zu finden. Eine solche Auslegung halte ich für verfehlt. Denn abgesehen davon, dass das so nicht im Text steht, waren die Menschen, selbst wenn man historisch-kritisch und evolutionär denkt, nie so naiv, auf eine derartige Idee zu kommen.
Dass Genesis 1 in der Zeit der babylonischen Gefangenschaft als „Antwort auf den Kulturdruck der Babylonier“ entstanden sei, gilt bekanntlich als Standard unter den Theologen, aber die Begründung dafür war sehr kurz. Es stellt sich hier die Frage, wie unter solchen Umständen den zuvor exegetisch begründeten Aussagen ein Realitätsgehalt zukommen kann. Woher hatten die Autoren ihr Wissen? Hatten sie es schon mitgebracht und wollten sie es nun aufschreiben? Oder ist es ihnen erst in Babylon in den Sinn gekommen? Wie kamen sie dann überhaupt darauf? Und mit welcher Begründung kann es als realistisch und glaubwürdig angesehen werden? Konkret: Was kann z. B. der ursprüngliche Tierfrieden überhaupt bedeuten, wenn es diesen in Wirklichkeit nie gab? Oder: Was soll es bedeuten, dass der Mensch – zum Bilde Gottes geschaffen – Gottes „Standbild“ (Stellvertreter) auf der Erde ist, wenn der Mensch in Wirklichkeit evolutionär bedingt ein höchst unvollkommenes Wesen ist, das einer solchen Aufgabe niemals gewachsen sein kann? Die zuvor vorgetragene Auslegung von Gen 1 ist damit unglaubwürdig. Wenn diese Texte also gar keinen (oder nur einen unbedeutenden) historischen Realitätsgehalt haben, verlieren sie auch ihren tieferen und existentiellen Sinn. Hier liegt das Hauptproblem neuzeitlicher Hermeneutik: Was das Alte und Neue Testament als wirkliches Handeln und Reden Gottes bezeugt (beginnend mit der Schöpfung), was in der Kirche so geglaubt wurde, sieht man heute lediglich als subjektive Vorstellung früherer Theologen bzw. religiöser Gruppen. Der hermeneutische Versuch, man solle die Schöpfungstexte als Zukunftstexte lesen, erscheint mir sehr merkwürdig, nicht textgemäß und ohne exegetische Begründung.
Schließlich muss man fragen: Wenn man schon argumentiert, Gen 1 sei (auch) Abgrenzung gegen babylonische Vorstellungen, warum gehört dann das astral gesteuerte Weltzeitalter-Schema mit seinen zyklischen Feuer- und Wasserkatastrophen nicht zu den Abgrenzungen? Denn gerade hier liegt doch ein markanter Unterschied der Zeitvorstellungen in Gen 1 zu damals zeitgenössischen Vorstellungen der israelitischen Umwelt im Exil vor. Was ist also der Auslegungsschlüssel? Ist es nicht doch das moderne Weltbild?
Oemings Bemerkung, die „fundamentalistische“ Bibelauslegung sei aus Ängsten gespeist, könnte man insbesondere allen Entflechtungslösungen vorhalten, die sich – etwas provozierend gesagt – vor lauter Angst auf sicheres, unangreifbares Terrain zurückgezogen haben, indem sie die Beziehung zur biblisch bezeugten Historie und damit zur Wirklichkeit des Handelns und Redens Gottes faktisch gekappt haben. Wenn aber, wie Oeming feststellte, das von ihm vorgetragene Schöpfungsverständnis unabhängig von den Ergebnissen der Naturwissenschaft sei, dann stellt sich die Frage, was überhaupt der Inhalt der Schöpfungslehre ist und ob sie einen Bezug zur Realität hat. Die Umweltproblematik kann ja trotz ihrer Wichtigkeit nicht der einzige Bezug sein. Es drängt sich der Eindruck auf, als würden die exegetisch ermittelten Inhalte aufrechterhalten, aber die sie begründenden faktischen Grundlagen bestritten. Im Bild gesprochen: Man behauptet, ein Mittagessen zu genießen, bestreitet aber gleichzeitig dessen Existenz.
Die Schlussfolgerungen gingen nach langer Rede in wenigen Minuten am Ende von Oemings Vortrag fast unter. Kreationisten nähmen biblische Texte selektiv wahr, so Oeming, die Begründung war für mich nicht mehr erkennbar Dagegen stellte sich bei Oemings Auslegung von Gen 2 umgekehrt die Frage, ob er Dinge in die Texte hineinliest. Leider blieb fast keine Zeit mehr für kritische Rückfragen.
Die Religionspädagogin Astrid Dinter analysierte im ersten Teil Ihres Vortrags die Argumentation in Dawkins’ Buch „Der Gotteswahn“ und diagnostizierte ein monistisches, einliniges naturwissenschaftliches Denken, das alles erklären wolle (die Tagungsteilnehmer hatten das zuvor live im Eingangsreferat von Thomas Junker erleben können). Wie einige andere Redner ging auch sie auf ID ein und kritisierte an diesen Ansatz ähnlich wie an Dawkins’ Buch eine „einlinige Argumentation“ und eine monistische Weltsicht. Leider gab es in keinem der Beiträge eine zusammenhängende Erklärung des ID-Ansatzes (ich hatte auch kaum Zeit dazu in meinem Vortrag). Wenn das Tagungsthema „Gott als Designer?“ heißt, wäre es sinnvoll gewesen, einen eigenen Grundsatzvortrag eines ID-Sympathisanten einzuplanen, in dem die Argumentationsweise im Zusammenhang dargestellt wird. Wichtig war hier die oben erwähnte Unterscheidung einer schwachen und schwachen Form von ID. Auf die schwache Form (die der Position der SG Wort und Wissen nahe kommt), trifft die o. g. Kritik von Frau Dinter nicht zu, da in dieser Version die Grenzen der Naturwissenschaft berücksichtigt werden. Hier wurde wie so oft bei „ID“ das Kind mit dem Bade ausgeschüttet. Im zweiten Teil thematisierte Frau Dinter religionspädagogische Aspekte des Themenfelds Glaube und Naturwissenschaft. Sie betonte die Wichtigkeit einer Verhältnisbestimmung von Theologie und Naturwissenschaft in der Pädagogik, wobei altersgemäß unterschiedlich vorgegangen werden muss. Gelinge hier nicht frühzeitig eine Synthese, drohe später ein Abbrechen der religiösen Weltsicht oder eine Ablehnung der Naturwissenschaft. Die SG Wort und Wissen teilt dieses Grundanliegen und möchte ebenfalls gerade diesen Abbruch sowohl zu Glaubensinhalten als auch zur Naturwissenschaft verhindern, doch die Vorstellungen zur konkreten Umsetzung dieses Anliegens gehen freilich in eine deutlich andere Richtung. Die historisch-kritische Ausdünnung der historischen und naturkundlichen Bezüge der Glaubensinhalte und des biblischen Zeugnisses (vgl. Oemings Vortrag) halten wir für eine schlechte Basis einer In-Bezug-Setzung von Glaube und Naturwissenschaft. Dass viele Jugendliche sich vom christlichen Glauben abwenden, könnte auch einer gefühlten Belanglosigkeit geschuldet sein, wenn die Bezüge zur realen Welt weitgehend gekappt worden sind.
Autor dieser News: Studiengemeinschaft Wort und Wissen
© 2008, http://www.genesisnet.info/schoepfung_evolution/n115.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
17.05.08 Ausführlicher Artikel zu „Evo-Devo“
Im Newsbeitrag „Evo-Devo: Schlüssel für Makroevolution?“, https://www.genesisnet.info/index.php?News=99 wurde bereits auf den in den letzten Jahren immer stärker aufkommenden Evo-Devo-Ansatz in der Evolutionsforschung und einen Genesisnet-Artikel zu diesem Thema hingewiesen. „Evo-Devo“ steht für „evolutionary developmental biology“, also für eine Synthese von Erkenntnissen der Erforschung der Entwicklungsbiologie (Ontogenese (= Individualentwicklung) der Lebewesen) und der Erforschung der Evolutionsmechanismen. Viele Evo-Devo-Forscher halten die verschiedenen Versionen der Synthetischen Evolutionstheorie für unzureichend, um die Entstehung des Neuen in der Evolution zu erklären und suchen nach neuen Antworten. Die Hoffnung ist, dass durch das verbesserte Verständnis der Vorgänge während der Ontogenese eine Erklärung für Makroevolution gefunden werden kann (zu Makroevolution siehe „Mikro- und Makroevolution“, https://www.genesisnet.info/index.php?Artikel=41223&Sprache=de&l=1).
Mittlerweile ist zu diesem aktuellen Forschungsfeld ein ausführlicher Artikel verfügbar, der als PDF heruntergeladen werden kann: „Evo-Devo“, https://www.genesisnet.info/pdfs/Evo-Devo.pdf (ca. 1,3 MB, 45 Seiten, 21 Abbildungen).
Der neue Artikel hat zum Teil (vor allem in den Anmerkungen) den Charakter einer Stoffsammlung, da viele Beispiele und Hypothesen nicht ausführlich diskutiert werden können. Um den Text lesbar zu halten, wurden viele Zitate, Beispiele und andere Zusatzinformationen in einem größeren Anmerkungsteil zusammengestellt. Die Anmerkungen sollen auch weitere Belege für im Haupttext aufgestellte Behauptungen liefern. Eine kurze Zusammenfassung bietet der eingangs verlinkte Newsbeitrag.
Autor dieser News: Studiengemeinschaft Wort und Wissen
© 2008, http://www.genesisnet.info/schoepfung_evolution/n111.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
15.01.08 Gotteswahn oder Selektionswahn?
Richard Dawkins macht mit seinem Buch „Der Gotteswahn“ (Ullstein-Verlag, 2007) viel von sich reden. Mich interessierte besonders das 4. Kapitel „Warum es mir ziemlicher Sicherheit keinen Gott gibt“, da es dort um Evolution und Schöpfung geht. Außerdem trägt Dawkins in diesem Kapitel nach eigener Aussage „die zentrale Argumentation“ seines Buches vor (222). Die Ausführungen in diesem Kapitel sind offenkundig für unkundige und / oder leichtgläubige Leser geschrieben. Selten habe ich eine dermaßen oberflächliche Abhandlung über Evolutionsmechanismen gelesen.
Dawkins möchte nachweisen, dass „die Darwin’sche natürliche Selektion die einzige bekannte Antwort auf die ansonsten unlösbare Frage“ ist, „woher die Informationen stammen“ (157). Natürliche Selektion zerstöre in der Biologie die „Illusion der gezielten Gestaltung“ (163) und sei eine Erklärung „für die Gesamtheit alles Lebendigen“ (160). Hier wird also ein maximaler Erklärungsanspruch erhoben. Wie wird er begründet?
Erstens: Eine „intelligente Gestaltung“ würde die Frage nach der Entstehung des Gestalters aufwerfen. Das führe in einen unendlichen Regress und sei daher keine Alternative. Zweifellos stehen wir hier vor eine Grenze unseres Verstehens, aber das ist kein Argument gegen die Annahme eines Gestalters. Wir lehnen die Erklärung durch einen Gestalter ja auch nicht ab, wenn wir eine Erklärung für die Form eines schön gearbeiteten Faustkeils suchen.
Die zweite Begründung für die Allmacht der Selektion lautet im Bild gesprochen: Selektion müsse nicht auf einmal einen Berg über eine Steilwand erklimmen (d. h. eine neue Struktur hervorbringen), sondern könne das in kleinen Schritten auf einem flacheren Hang tun. Hier müsste jetzt jedoch eine genauere Betrachtung dazu ansetzen, wie groß die einzelnen Schritte sind, die die Selektion immer noch bewältigen muss, wenn der Berg schrittweise erklommen werden soll. Hierzu verweist Dawkins lediglich auf Formenreihen wie die oft zitierten Augentypen. Schon Darwin habe „mühelos“ erklären können, „wie sich das Auge in der Evolution ganz allmählich entwickelt hat“ (171). Formenreihen begründen aber zum einen keine Mechanismen (um die es hier gerade geht), zum anderen liegt das Problem im Detail: was muss sich von Augentyp zu Augentyp verändern und gibt es dafür plausible Modelle? (siehe dazu http://www.si-journal.de/index2.php?artikel=jg13/heft1/sij131-1.html) Nichts davon diskutiert Dawkins. Stattdessen Polemik gegen Kreationisten und unzutreffende Feststellungen über sie: „Die Kreationisten dagegen sind blind für alles mit Ausnahme der steil aufragenden Felswand auf der Vorderseite“ (170); die „Leistung der Akkumulation“ begriffen sie nicht. Nein, Dawkins kennt nicht ihre Argumente, die es auch gegen den Flachaufstieg, gegen eine Akkumulation gibt, oder er übergeht sie (siehe o. g. Link). Statt sich mit diesen Argumenten auseinanderzusetzen, macht er die Kreationisten lächerlich. Dawkins Empörung über die Religion, die sich mit Nichtwissen zufrieden gebe und dies auch noch als Tugend betrachte (S. 175), fällt auf ihn selbst zurück.
Dieses Nichtwissen Dawkins’ fällt besonders bei der Besprechung des Nano-Rotationsmotors von Bakterien auf, und zwar in mehrfacher Hinsicht: Er schreibt, der Biochemiker Michael Behe (bekannt durch das Buch „Darwin’s Black Box“, siehe Kurzrezension unter https://www.wort-und-wissen.org/rezension/darwins-black-box-biochemische-einwaende-gegen-die-evolutionstheorie/) würde ohne ein Wort der Begründung behaupten, der Bakterienmotor sei nichtreduzierbar komplex (S. 183). Das ist schon eine sehr dreiste Unterstellung. Dann unterstellt er weiter, Behe halte keines der Einzelteile des Motors für sich alleine für nützlich. Das ist falsch; Behe hat im Gegenteil oft genug gesagt, dass Einzelteile eines nichtreduzierbar komplexen Systems eine andere Funktion als das System selbst ausüben können. Und schließlich verweist er auf den Typ-III-Sekretionsapparat, der eine Vorstufe des Bakterienmotors sein solle. Doch das widerlegt Behes Argumentation nicht, da er die Möglichkeit von Vorstufen mit anderer Funktion ausdrücklich berücksichtigt, und vor allem weil beim hypothetischen Übergang vom Sekretionsapparat zum Motor zahlreiche zusätzliche Proteine benötigt werden (Details s. Junker & Scherer, Evolution – ein kritisches Lehrbuch, 159ff.); ein Übergang ist völlig unverstanden. Dazu kommt, dass es – Evolution vorausgesetzt – starke Argumente dafür gibt, dass der Sekretionsapparat vom Motor abzuleiten ist und nicht umgekehrt (siehe dazu den betreffenden Abschnitt in: Nichtreduzierbare Komplexität, https://www.genesisnet.info/pdfs/Irreduzible_Komplexitaet.pdf). Von allen diesen Argumenten erfährt der Leser von Dawkins nichts. Man muss es hier sagen: Das ist in hohem Maße unredlich.
Selektion erweitere unser Bewusstsein, meint Dawkins (160), „wie aus einfachen Anfängen ohne absichtliche Lenkung organisierte Komplexität entsteht.“ Außer Formenreihen und Optimierungsbeispielen bietet Dawkins jedoch nur mehrfache Wiederholungen des schon Gesagten, um den Funken dieser Bewusstseinserweiterung überspringen zu lassen, und versucht darüber hinaus wieder und wieder mit Polemik und Verächtlichmachen Andersdenkender nachzuhelfen. Man gewinnt den Eindruck, Dawkins wolle dem Leser durch ständige Wiederholungen seine Weltsicht eintrichtern.
Mit billiger Argumentation geht es weiter: „So unwahrscheinlich die Entstehung des Lebens gewesen sein mag, wir wissen, dass sie sich auf der Erde abgespielt hat, denn wir sind hier!“ (192f.) Dawkins schätzt die Chance, dass Leben durch natürliche Vorgänge auf unserer Erde entstehen könnte, auf 1 : 1 Milliarde. Eine seriöse Begründung für diesen sehr optimistischen Wert liefert er nicht. Gegenargumente werden nicht angesprochen, stattdessen meint er, es gebe im Universum wohl so viele erdähnliche Planeten, dass es klar sei, dass auf manchen auch Leben entstehen konnte. Dem „Intelligent Design“ sei damit der Todesstoß versetzt (195). In Wirklichkeit argumentiert Dawkins hier auf der Basis von Nichtwissen. Denn weder ist außer unserer Erde ein einziger erdähnlicher Planet bekannt, noch kann man eine begründete quantitative Abschätzung darüber machen, wie wahrscheinlich die Entstehung des Lebens ist. Dawkins macht hier aus Nichtwissen ein gesichertes Wissen. Eine Erklärung für dieses unhaltbare Vorgehen könnte sein Bekenntnis sein, dass Chemie nicht sein Gebiet sei (192). Dawkins muss wirklich auf uninformierte und leichtgläubige Leser hoffen.
Wie schon angesprochen verbindet Dawkins die mangelhafte bis ungenügende Argumentation mit ätzender Polemik. Schöpfungsgläubigen unterstellt er in Unkenntnis über die Geschichte der Naturwissenschaft, sie wollten nicht forschen (176, 185), sie würden Lücken „automatisch“ durch Gott füllen (178) und stellt Schöpfungsgläubige mehr oder weniger offen, teilweise subtil als Dummköpfe dar (z. B. 179), und der Schritt zur Verspottung Gottes ist dann nicht mehr weit (164). Ob er damit „bekehren“ kann, was er ausdrücklich anstrebt (160)?
A propos „bekehren“: Ein Stück weit bekehren muss er auch einige seiner Evolutionsbiologen-Kollegen, die der natürlichen Selektion nicht die Allmacht zugestehen, die „Gesamtheit alles Lebendigen“ (160) zu erklären. Man denke hier an die Evo-Devo-Forscher, deren Programm unter anderem gerade dadurch motiviert ist, dass Makroevolution durch die Selektionstheorie eben nicht erklärt ist (siehe dazu Evo-Devo, https://www.genesisnet.info/index.php?Artikel=41266&Sprache=de&l=2 und Mikroevolution, Makroevolution und „ID“, https://www.genesisnet.info/index.php?News=93; diese Forscher bezweifeln freilich nicht, dass es Makroevolution gegeben hat).
Noch zwei interessante Sätze sind mir aufgefallen: „Wer ein Argument als ‘neunzehntes Jahrhundert’ bezeichnet, hat damit noch nicht erklärt, was falsch daran ist“ (220). Dawkins bezieht das auf Darwins Vorstellungen; man kann es natürlich auch auf anderes beziehen. Und der zweite: „Erst Darwin lieferte den unentbehrlichen Kran, an dem Hume natürlich seine helle Freude gehabt hätte“ (221f.). Der Kran steht für die natürliche Selektion, die nach und nach etwas hochhieven kann (s.o.). Warum hätte dieser Kran David Hume entzückt? Der Grund: Hume wird nachgesagt, er hätte das Design-Argument widerlegt1; dem widerspricht Dawkins an dieser Stelle, mit dem Hinweis, dass Hume den Kran eben noch nicht kannte. Also konnte er den „Himmelshaken“ (wie Dawkins die direkte Gestaltung durch einen Schöpfer nennt), nicht aufgeben. „Erst Darwin“ konnte das. Wenn sich das aber als voreilig erwiesen hat, dann sind alte Ideen, die noch älter als das 19. Jahrhundert sind, immer noch topaktuell!
Anmerkung
1 Was Hume genau widerlegt hat und was nicht, müsste eigens genauer beleuchtet werden, was den Rahmen dieses Newsbeitrags sprengen würde.
Autor dieser News: Reinhard Junker
© 2008, http://www.genesisnet.info/schoepfung_evolution/n104.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
19.11.07 Evo-Devo: Schlüssel für Makroevolution?
„Evo-Devo“ ist ein relativ neues Schlagwort in der Diskussion um Evolutionsmechanismen, speziell wenn es um die Aufklärung der Mechanismen für Makroevolution geht. Dass diese Mechanismen nicht oder nicht genügend bekannt sind, wird auch von manchen Evolutionsbiologen eingeräumt (vgl. Mikroevolution, Makroevolution und „ID“, https://www.genesisnet.info/index.php?News=93).
In einem neuen Artikel (Evo-Devo, https://www.genesisnet.info/index.php?Artikel=41266&Sprache=de&l=2) werden die wichtigsten Befunde zusammengestellt, die zur Entwicklung des Evo-Devo-Ansatzes geführt haben. Es folgt eine Darstellung der Grundzüge von Evo-Devo-Hypothesen, die anschließend kritisch auf ihre Leistungsfähigkeit hin beurteilt werden. Im Einzelnen:
- Was bedeutet Evo-De vo?
- Anlässe für Evo-Devo
- Wie funktioniert Evo-Devo?
- Kritik an Evo-Devo
- Wo steht Evo-Devo?
„Evo-Devo“ steht für „evolutionary develomental biology“ und mithin für eine Synthese von Erkenntnissen der Erforschung der Entwicklungsbiologie (Ontogenesen, d. h. Individualentwicklung, der Lebewesen) und der kausalen Evolutionsforschung. Mit dem Evo-Devo-Ansatz ist die Hoffnung verbunden, durch das Verständnis der Vorgänge während der Ontogenese einen Schlüssel für ein besseres bzw. vollständiges Verständnis evolutionärer Prozesse zu gewinnen, insbesondere für die Erklärung von Makroevolution.
Kennzeichnend für Evo-Devo-Forscher ist die Auffassung, dass der Neodarwinismus bzw. die Synthetische Evolutionstheorie („Modern Synthesis“) den evolutionären Wandel nicht vollständig erklären kann. Manche Evo-Devo-Forscher formulieren die Defizite des Neodarwinismus schärfer und halten wesentliche Fragen der Entstehung evolutionärer Neuheiten für ungeklärt (zu „evolutionären Neuheiten“ vgl. Mikro- und Makroevolution, https://www.genesisnet.info/index.php?Artikel=41223&Sprache=de&l=1).
Reinhard Junker zeigt in seiner Analyse, dass auch der Evo-Devo-Ansatz bislang in wichtigen Fragen keine evolutionstheoretisch plausiblen Antworten geben kann. Die wichtigsten Begründungen in Kurzform:
- Die Mechanismenfrage bleibt ohne überzeugende Antwort. Evo-Devo-Forscher sprechen von Neuprogrammierung, Rekrutierung, Kooption oder Flickschusterei; die vorgeschlagenen Vorgänge sind aber hypothetisch und ohne Lenkung (die es im Evolutionsprozess nicht gibt) unplausibel.
- Häufig wird vergleichend-biologisch argumentiert. Vergleichende Biologie begründet, aber keinerlei Mechanismen und begründet nicht einmal Evolution (vgl. Ähnlichkeiten in der Morphologie und Anatomie.
- Die Triebfeder für Neuverwendung von Steuergenen in ganz verschiedenen Zusammenhängen ist unklar.
- „Konstruktionszwänge“, die einerseits eine Lenkung der Evolution ermöglichen sollen, verhindern andererseits Veränderungen in Richtung neuer Konstruktionen.
- Neuprogrammierung ist nicht alles. Die Neuprogrammierung von Regulationsgenen und anderen sog. „tool-kit“-Genen gilt als wichtiger (hypothetischer) Evo-Devo-Mechanismus. Doch die Frage, woher die sehr verschiedenen Gene bzw. Genfamilien kommen, wird dadurch nicht beantwortet.
- Schließlich fällt die Verwendung von „Schöpfungsvokabular“ auf.
Diese und weitere Kritikpunkte werden in „Evo-Devo“ ausführlich erläutert: https://www.genesisnet.info/index.php?Artikel=41266&Sprache=de&l=2.
Autor dieser News: Studiengemeinschaft Wort und Wissen
© 2007, http://www.genesisnet.info/schoepfung_evolution/n99.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
07.04.06 Mikroevolution und Makroevolution: fließende oder scharfe Grenze?
In den Kontroversen um die Plausibilität einer allgemeinen Evolution aller Lebewesen hat das Begriffspaar „Mikroevolution – Makroevolution“ eine besondere Bedeutung. Die inhaltliche Bestimmung ist allerdings in den biologischen Publikationen sehr uneinheitlich, und nicht selten wird diese Begriffsdifferenzierung mindestens in der Frage der Mechanismen der Evolution für unnötig erachtet. Die Verwendung dieser Begriffe wäre demnach nur sinnvoll, um graduelle Unterschiede im Ausmaß von evolutionären Veränderungen zum Ausdruck zu bringen; es gebe letztlich nur einen Evolutionsmechanismus und dabei keine qualitativen Unterschiede.
Damit ist die Kontroverse um das Begriffspaar Mikroevolution – Makroevolution eröffnet. Die beiden Begriffe werden oft verwendet, wenn es um die Frage geht, wie gut die Theorie von einer allgemeinen Evolution aller Lebewesen begründet ist und ob die Mechanismen für eine umfassende Evolution der Lebewesen als aufgeklärt gelten können. Diese Frage wird innerhalb der Evolutionsbiologie auch heute durchaus kontrovers diskutiert.
In einem neuen Beitrag über diese beiden Begriffe („Mikro- und Makroevolution“, https://www.genesisnet.info/index.php?Artikel=41223&Sprache=de&l=2) gibt Reinhard Junker zunächst (in Anlehnung an einen Internetbeitrag von Thomas Waschke) einen kurzen Überblick über verschiedene Vorschläge einer inhaltlichen Bestimmung des Begriffs „Makroevolution“. Er plädiert dafür, eine Unterscheidung zwischen Mikroevolution und Makroevolution an der Qualität der Veränderungen der Lebewesen und an entsprechenden unterschiedlichen Fragestellungen und Erklärungsproblemen (kurz: Optimierung oder Konstruktion?) festzumachen. Nur dann könne die Frage nach der Entstehung des Neuen in der Biologie adäquat formuliert und darauf Forschung aufgebaut werden. Kurz gefasst:
Mikroevolution: Wie werden vorhandene Konstruktionen der Lebewesen optimiert?
Makroevolution: Wie entstehen Konstruktionen erstmals – de novo?
Im Artikel „Mikro- und Makroevolution“ (https://www.genesisnet.info/index.php?Artikel=41223&Sprache=de&l=2) werden des weiteren verschiedene Argumente für eine Gleichsetzung von Mikroevolution und Makroevolution besprochen.
Es wird deutlich, dass es in der Frage nach der Unterscheidung von Mikroevolution und Makroevolution letztlich um die Reichweite der bekannten Evolutionsfaktoren geht.
Ein ausführlicherer Beitrag zu diesem Thema ist als PDF herunterladbar: „Mikro- und Makroevolution“, https://www.genesisnet.info/index.php?Artikel=41223&Sprache=de&l=2. Dieser Artikel enthält zusätzlich einen Abschnitt, in welchem einige wichtige neuere Erklärungsansätze für Makroevolution vorgestellt und kritisch beleuchtet werden. Im Anhang ist außerdem eine ausführliche Rezension des Buches „The Plausibility of Life. Resolving Darwin’s Dilemma“ von Marc Kirschner und John Gerhart aufgenommen worden. Kirschner & Gerhart präsentieren in diesem Ende 2005 erschienenen Buch Ideen zur Lösung der Frage, wie evolutionäre Neuheiten (= „Darwins Dilemma“) entstanden sind (wie also Makroevolution funktioniert). In der Rezension wird folgendes Fazit gezogen: „‘The Plausibility of Life’ ist ein lesenswertes Buch, es bietet viele faszinierende Erkenntnisse aus der Forschung der letzten Jahre. Wie man diese Erkenntnisse im Hinblick auf die Ursprungsfragen interpretiert, ist zu einem gewissen Teil Sache der Perspektive. Kirschner & Gerhart bieten zweifellos interessante Aspekte zur Variations- und Anpassungsfähigkeit der Lebewesen. Dass die von ihnen vorgestellten Befunde mehr hergeben und ‚Darwins Dilemma‘ lösen, kann aus den genannten Gründen jedoch ernsthaft bezweifelt werden.“ Zu einer ähnlichen Schlussfolgerung gelangen zwei Evolutionsbiologen in ihren Rezensionen: „Kirschner and Gerhart do not present any detailed examples of how the properties of developmental systems have actually contributed to the evolution of a major evolutionary novelty“ (Charlesworth B (2005) On the origins of novelty and variation. Science 310, 1619-1620). „But with its sometimes troubling limitations, the book falls short of the major new theory that the authors promise in their introduction“ (Erwin D (2005) A variable look at evolution. Cell 123, 177-179).
In „The Plausibility of Life“ geht es auf einigen Seiten auch um „Intelligent Design“ und um „Irreduzible Komplexität“. Auch darauf wird in der Rezension eingegangen.
Autor dieser News: Studiengemeinschaft Wort und Wissen
© 2006, http://www.genesisnet.info/schoepfung_evolution/n62.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
02.01.06 Charles Darwin, der Held des Jahres 2005
Das renommierte Wissenschaftsjournal Science feiert in der Ausgabe vom 23. Dezember Charles Darwin als Helden des Jahres 2005, weil jüngste Daten die auf ihn zurückgehende Evolutionstheorie untermauert hätten. Sie werden sogar als „Durchbruch des Jahres“ gefeiert. Die im Artikel von Elizabeth Culotta und Elizabeth Pennisi (Science 310, 1878-1879) genannten Begründungen wirken allerdings sehr dürftig. So wird beispielsweise die Entschlüsselung des Affengenoms genannt. Die 1% Unterschiede zum Menschen machen 40 Millionen Evolutionsschritte aus, schreiben die Autorinnen. Wie diese erfolgt sind, ist jedoch nicht Gegenstand der betreffenden Untersuchungen, sondern die Sequenzierung des Erbguts des Schimpansen und dessen Vergleich mit dem Genom des Menschen. Mit Evolution an sich hat das nichts zu tun. Vielmehr wird hier ein Argument zugrundegelegt, das in der öffentlichen Debatte um Evolution gewöhnlich unkritisch verwendet wird: das Argument nämlich, man könne aus Ähnlichkeiten auf Evolution schließen und Abstammungsverhältnisse bestimmen. Die evolutionäre Interpretation von Ähnlichkeiten ist aber nicht stichhaltig (siehe Ähnlichkeiten in der Morphologie und Anatomie, https://www.genesisnet.info/index.php?Artikel=41301&Sprache=de&l=1 und Molekularbiologie, https://www.genesisnet.info/index.php?Artikel=41302&Sprache=de&l=2; insbesondere den Expertenteil von Ähnlichkeiten in der Morphologie und Anatomie, https://www.genesisnet.info/index.php?Artikel=41301&Sprache=de&l=2). Ähnlichkeiten lassen sich zudem auch im Rahmen einer Schöpfungsvorstellung deuten (Mosaikformen als Grundtypen und Baukastensysteme, https://www.genesisnet.info/index.php?Artikel=1261&Sprache=de&l=1). Das gilt auch für molekulare Ähnlichkeiten. Genomdaten an sich geben entgegen der Behauptung von Science keine Hinweise auf die Vorgänge, die zum Umbau von Mikroben bis hin zu den Primaten geführt haben. Auch darüber, wie die Unterschiede zwischen Mensch und Schimpanse entstanden sind, gibt es nur allgemeine theoretische Modelle, die bislang kaum prüfbar sind. (Zu diesen Mechanismenfragen siehe zum Beispiel Gene tinkering, https://www.genesisnet.info/index.php?Artikel=41264&Sprache=de&l=1 und Homeobox-Gene und Evolution, https://www.genesisnet.info/index.php?Artikel=41265&Sprache=de&l=1.) Zudem sollte es sich mittlerweile herumgesprochen haben, dass die genetischen Unterschiede bei weitem nicht alleine die Unterschiede zwischen den mannigfaltigen Formen des Lebens ausmachen. Wo die biologischen Unterschiede zwischen Mensch und Schimpanse liegen, ist ein spannendes Forschungsthema. Und ob auf dieser Ebene die entscheidenden Unterschiede überhaupt festgemacht werden können, ist vorerst nicht ausgemacht. Was gibt dem Menschen die Fähigkeit, eine Chopin-Ballade auf dem Klavier zu spielen, Ballett zu tanzen oder den genetischen Code zu entschlüsseln – Fähigkeiten, von denen Schimpansen höchstwahrscheinlich nicht einmal träumen?
Weiter werden im Science-Artikel eine neue Klassifikation der Mikroorganismen und die Entdeckung eines Dinosaurier-Embryos als Beispiele erwähnt. Inwiefern diese Erkenntnisse Belege für Darwins Evolutionstheorie sein sollen, ist nicht ersichtlich. Hier werden wie im Falle der Entschlüsselung des Affengenoms die Befunde ohne weitere Begründung als Bestätigungen für die Evolutionstheorie reklamiert.
Der Science-Artikel ist mit „Evolution in action“ überschrieben. Auch dafür werden Beispiele genannt, doch es handelt sich ausschließlich um mikroevolutive Vorgänge wie Artbildung (vgl. Artbildung, https://www.genesisnet.info/index.php?Artikel=41244&Sprache=de&l=1). Die Aufspaltung einer Art ist jedoch nicht nur ein Baustein im Gebäude der Evolutionstheorie, sondern auch ein wichtiger Bestandteil der Grundtypenbiologie der Schöpfungslehre (vgl. Heutige Grundtypen, https://www.genesisnet.info/index.php?Artikel=1241&Sprache=de&l=1). Auch in dessen Theorierahmen werden Variationsmechanismen vorhergesagt. Nach dem Konzept der polyvalenten Stammformen (Genetisch polyvalente Stammformen von Grundtypen, https://www.genesisnet.info/index.php?Artikel=1244&Sprache=de&l=1) sind Anpassungs- und Artbildungsvorgänge in schnellem Tempo zu erwarten, weil von einem großen Variationspotential und programmierter Variabilität ausgegangen wird. Das von den Science-Autorinnen genannte Beispiel der Divergenz der Mönchsgrasmücke und ähnliche Fälle sind daher schöne Beispiele zur Veranschaulichung des Grundtypenkonzepts (vgl. R. Junker & S. Scherer, Evolution – ein kritisches Lehrbuch, Gießen 2001, Seite 293).
Weiter werden Veränderungen an Stichlingen angeführt. Im Meer lebende (marine) Stichlinge tragen eine Panzerung aus Knochenplatten, die Süßwasserarten fehlt. Hauptverantwortlich dafür sind Unterschiede in einem bestimmten Gen (Ectodysplasin-Gen). Dieses Gen existiert in zwei Varianten (Allelen). Das Allel „ungepanzert“ kommt mit geringer Häufigkeit auch in marinen Stichlingen vor. Die fehlende Panzerung der Süßwasser-Stichlinge, die aus marinen Arten entstanden sind, ist daher ein Selektionsereignis (N. Winkler: Stichlinge: Evolution oder Allelfrequenzverschiebung? Studium Integrale Journal 12 (2005), 76-77), aus dem keine Erkenntnisse über die ursprüngliche Entstehung der Knochenplatten gewonnen werden können. Auch dieses Beispiel passt gut ins Konzept polyvalenter Stammformen im Rahmen der Grundtypenbiologie und ist kein Beleg für Makroevolution.
Dass sich fernerhin mehr und mehr herausstellt, dass die ehemalige „Junk-DNA“ – der vermeintliche genetische Abfall der Evolution – doch kein Müll ist, ist sicher ebenfalls kein Grund, die Erklärungskraft der Evolutionstheorie zu feiern. Oft genug wurde unter Verweis auf die „Nonsense“-Gene über Schöpfungslehre gespottet. Nun erweist sich deren Vorhersage, dass die nicht-codierenden Teile des Genoms sinnvolle Funktionen erfüllen, mehr und mehr als Treffer.
Vor dem Hintergrund der Begründungen für den „Durchbruch des Jahres“ wirkt der Satz der Science-Autorinnen „At some level every discovery in biology and medicine rests on it [der Evolutionstheorie]“ fast wie ein Glaubensbekenntnis. Es wäre eine spannende Sache, den Nachweis für diese Behauptung zu erbringen.
Autor dieser News: Reinhard Junker
© 2006, http://www.genesisnet.info/schoepfung_evolution/n54.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
18.11.05 Was erklärt die Evolutionstheorie?
Am 11. Oktober 2005 referierte Reinhard Junker auf der Philosophischen Woche der Katholischen Akademie Bayern in München sowie am 1. November 2005 im Staatlichen Museum für Naturkunde in Görlitz einen Vortrag zum Thema „Was erklärt die Evolutionstheorie? Anfragen eines Kritikers“. Nach einer Erläuterung des Erklärungspotentials der Evolutionstheorie in drei ausgewählten Gebieten (Vergleichende Biologie, Paläontologie, kausale Evolutionsforschung) wurden im Hauptteil einige grundlegende Kritikpunkte zusammengestellt, mit denen das Evolutionsparadigma als Rahmenanschauung insgesamt in Frage gestellt wurde. Die Kritik betrifft sowohl die kausale als auch die historische Evolutionsforschung und kann daher zwei Problembereichen zugeordnet werden:
- Erklären die experimentell nachgewiesenen Variationsmechanismen die Entstehung neuer Baupläne? (kausale Evolutionsforschung)
- Sind die Belege für eine allgemeine Evolution der Lebewesen (im Sinne von Makroevolution) stichhaltig? (historische Evolutionsforschung)
Auf der Homepage der Studiengemeinschaft Wort und Wissen kann eine Langversion des Vortrags heruntergeladen werden: https://www.wort-und-wissen.org/wp-content/uploads/a03.pdf (PDF, 2,6 MB, umfasst 15 Seiten und enthält 14 Abbildungen)
Es werden folgende Schlussfolgerungen gezogen: Zahlreiche Daten der Biologie und Paläontologie können einigermaßen schlüssig im Rahmen der Evolutionstheorie verständlich gemacht werden, das heißt sie finden bei Vorgabe einer Makroevolution eine plausible Deutung.
Eine Reihe von Daten kann auch als Bestätigung von Vorhersagen gewertet werden, die aus der Evolutionstheorie resultieren, z. B. die Fossilabfolge (im Groben betrachtet) und die Ähnlichkeitshierarchie der Lebewesen.
Andererseits entsprechen zahlreiche Befunde, die regelmäßig auftreten, nicht den ursprünglichen evolutionstheoretischen Erwartungen. Dazu gehören:
- In der Vergleichenden Biologie: Das häufige Auftreten von Konvergenzen (insbesondere bei Komplexmerkmalen) und damit zusammenhängend die oft eher netz- als baumförmige Beziehung zwischen den Taxa sowie mangelnde Kongruenz zwischen verschiedenen Datensätzen.
- In der Paläontologie: Das plötzliche Auftreten von Fossilgruppen; dabei ist die Verschiedenartigkeit häufig bereits zu Beginn der Fossilüberlieferung erheblich, so dass auch abgeleitete Merkmale oder Taxa bereits früh in den jeweiligen Abfolgen fossil in Erscheinung treten.
- In der kausalen Evolutionsforschung: Eine Unterscheidung zwischen Mikro- und Makroevolution, die sich an der Qualität der Veränderungen der Lebewesen orientiert, ist unbedingt geboten. Makroevolution steht für die Entstehung neuer Konstruktionen, die nicht als Extrapolation von Variationsvorgängen wie Anpassungen, Spezialisierungen oder Optimierungen betrachtet werden kann. Für makroevolutive Veränderungen fehlen experimentelle Belege; dafür vorgeschlagene Mechanismen wie Evolution neuer funktionaler Gene durch Genduplikation, durch Exon Shuffling, durch Mutation von Hox-Genen oder durch Gen-Tinkering sind weitgehend hypothetisch. Insbesondere ist ungeklärt, wie Leben oder wenigstens die wichtigen Makromoleküle der Lebewesen erstmals unter unspezifischen Bedingungen einer hypothetischen frühen Erde entstanden sein könnten.
Die genannten Befunde falsifizieren die Evolutionstheorie zwar nicht, erforderten in der Vergangenheit aber wiederholt so schwerwiegende Änderungen im Theoriengebäude, dass man oft nicht mehr von Bestätigung von Vorhersagen sprechen kann, sondern nur von Deutungen im Nachhinein.
Von einer Erklärung des makroevolutiven Wandels im Sinne des Ableitens der empirischen Befunde aus allgemeinen Gesetzmäßigkeiten und spezifischen Randbedingungen kann nicht gesprochen werden. Lediglich für einige mikroevolutive Vorgänge trifft dies zu.
Auf http://www.evolutionsbiologen.de/PhilSemJ.pdf wird auf diesen Vortrag kritisch eingegangen; Interessierte können die dort gemachten Behauptungen mit den Ausführungen von Reinhard Junker direkt vergleichen. Da von der Akademie auch eine Tonaufzeichnung gemacht wurde, kann auch der Redetext zum Vergleich herangezogen werden.
Autor dieser News: Studiengemeinschaft Wort und Wissen
© 2005, http://www.genesisnet.info/schoepfung_evolution/n51.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
13.05.05 Schöpfungsvokabular bei Evolutionstheoretikern
Die Sprache verrät, was Sache ist. In Ursprungsfragen sei der Schöpfer „radikal ausgebootet“ worden, schrieb einmal der Verhaltensforscher Wolfgang Wickler. Doch der Schöpfer verfolgt die evolutionär orientierten Wissenschaftler weiter in vielen Formulierungen, die sie verwenden. Ein besonders eindrückliches Beispiel lieferte eine Meldung über die wahrscheinlich mehrmalige Entstehung der Gehörknöchelchen der Säugetiere.
Worum geht es? Aus einem neuen Fossilfund wird geschlossen, dass die säugertypischen Gehörknöchelchen Hammer, Amboss und Steigbügel mindestens zweimal unabhängig entstanden sein müssen. Dies ist evolutionstheoretisch sehr überraschend, da die Konfiguration der Gehörknöchelchen als eines der wichtigsten Schlüsselmerkmale der Säugetiere betrachtet wird. Die Bedeutung dieses Fundes für das Verständnis der Säugerevolution wird in der Herbstausgabe von Studium Integrale Journal (https://www.wort-und-wissen.org/publikationen/studium-integrale-journal/) diskutiert werden; hier interessiert uns jedoch ein anderer Aspekt. Antje Findeklee schreibt dazu in wissenschaft-online (www.wissenschaft-online.de/abo/ticker/77319; Hervorhebungen nicht im Original): „Hören ist eine verflixt komplizierte Angelegenheit. Und das dafür notwendige knöcherne wie häutige Inventar des Säugetierohr-Baukastens wirkt so hochspezialisiert, dass man nur staunen kann, wer wann im Laufe der Evolution dieses ausgeklügelte System entwickelt und seinen zahlreichen Nachfahren vom Schnabeltier bis zum Menschenkind hinterlassen hat.“ Das ist Schöpfungsvokabular: Ein Akteur („wer“) hat etwas „ausklügelt“ und benutzt einen Baukasten – nur, wer ist „er“?
Im Weiteren schreibt die Autorin: „… so etwas Raffiniertes würde selbst die höchst einfallsreiche Evolution wohl kaum zweimal erfinden, so verschlungen und überraschend deren Ideen auch manchmal sein konnten.“ Da hat sich also jemand eine raffinierte, überraschende Idee einfallen lassen!
Am erstaunlichsten sind jedoch die Schlusssätze. Nach der Feststellung, dass ein herausragendes Merkmal der Säugetiere seine einzigartige Stellung verliere, schreibt Findeklee: „Der Sache an sich tut das keinen Abbruch, im Gegenteil: Wie viel erstaunlicher ist nun die Erkenntnis, dass Mutter Natur den raffinierten Trick unabhängig ein zweites Mal entwickelte. Evolution sollte man einfach nie unterschätzen.“ So wird ein evolutionstheoretisch ursprünglich völlig unerwarteter Befund flugs ins Evolutionsgebäude eingebaut, indem man „der Evolution“ (die außerdem auch noch als „Meister“ und als „höchst einfallsreich“ bezeichnet wird) schöpferische Qualitäten zuspricht. Alles klar?
Richtig, das sind ja nur Metaphern! Man kann das auch anders sagen; allerdings wird es dann ziemlich kompliziert oder das betreffende Phänomen wird gar nicht angemessen beschrieben. Und meistens schleicht sich auch dann wieder Schöpfungsvokabular ein, z. B. wenn von „Selbstorganisation“ die Rede ist. Denn: Wer organisierte da?
Man könnte noch einwenden, dass dies die Sprache des Wissenschaftsjournalismus sei, doch dasselbe Vokabular wird auch in Fachartikeln verwendet. Offenbar drängt sich das Schöpfungsvokabular auf: Die beobachteten Phänomene lassen sich zwanglos beschreiben, wenn man ein durchdachtes Handeln eines Urhebers zugrunde legt (vgl. Einführung in „Intelligent-Design“, https://www.genesisnet.info/index.php?Artikel=1621&Sprache=de&l=1 und Kontroverse um „Intelligent-Design“, https://www.genesisnet.info/index.php?Artikel=1622&Sprache=de&l=1). Es gilt wohl auch für Biologen, was Paulus schreibt: Gottes Macht und Größe kann man an seinen Werken deutlich erkennen (Röm 1,18ff.).
Autor dieser News: Reinhard Junker
© 2005, http://www.genesisnet.info/schoepfung_evolution/n37.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/